Imprinting of a zein post-transcriptional regulator
--Sumita Chaudhuri and Joachim Messing
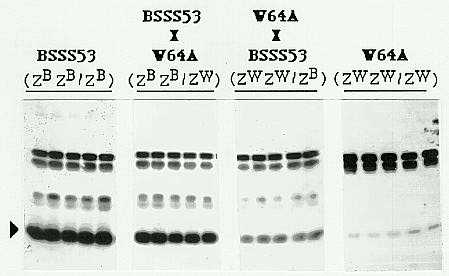
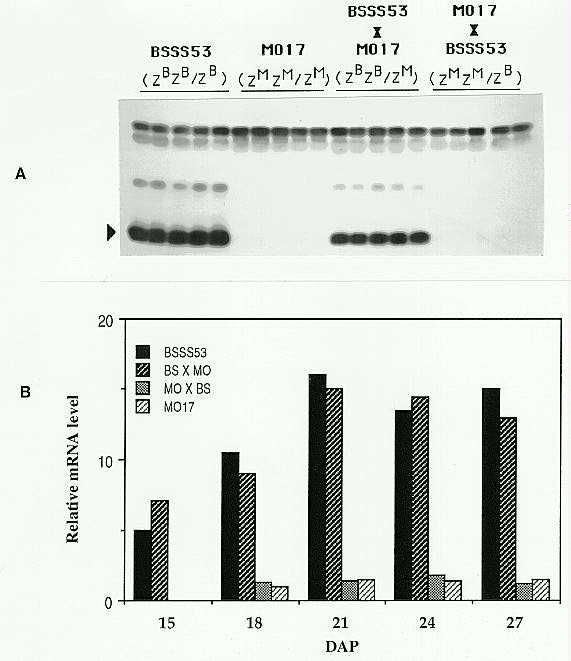
The level of expression of methionine-rich 10kD zein in maize endosperm is determined by a post-transcriptional regulator, Zpr10/22 (TAG 78:761, 1989; MGG 225:331, 1991). In mature endosperm of BSSS53, the level of 10kD zein is several times higher than in W64A, A654, A619 and Mo17 lines. Studies involving crosses of BSSS53 with W64A, A654 and A619 indicate that Zpr10/22 functions in a dose dependent manner in determining the 10kD zein level. For example, in reciprocal crosses of BSSS53 and W64A (Fig. 1), there is increased accumulation of 10kD zein with increasing doses of the Zpr10/22 allele from the BSSS53 parent. A similar dosage response is seen in the steady-state level of 10kD zein mRNA in immature endosperms of the same reciprocal crosses. However, in reciprocal crosses of BSSS53 and Mo17 the level of 10kD zein in the hybrids resembles that in the female parent instead of a dosage response (Fig. 2A). The steady-state 10kD zein mRNA level also shows maternal effect throughout endosperm development of the hybrids (Fig. 2B). This maternal effect cannot be due to maternal cytoplasmic or sporophytic factors since the 10kD zein level segregates with Zpr10/22 in backcross populations.
There are two possible explanations for this maternal effect. First, unequal gene dosage from the parents in the endosperm leads the allelic composition of Zpr10/22 to differ in the reciprocal hybrids (BSSS53 BSSS53/Mo17, Mo17 Mo17/BSSS53). Thus, at least two doses of the Mo17 allele may be necessary to downregulate the 10kD zein level. Ordinarily, two doses of the BSSS53 allele are not required to upregulate 10kD zein level. For instance, one dose is sufficient to upregulate the 10kD zein level in crosses with W64A.
Imprinting, i.e. unequal gene function following passage through male or female parent, is the other possible explanation for this maternal effect. As seen in crosses with W64A, the BSSS53 allele is functional irrespective of which parent transmits it, and is therefore not the allele imprinted. According to this model, therefore, the Mo17 allele is subject to imprinting so that it is a dominant downregulator of 10kD zein expression only when transmitted by the female parent.
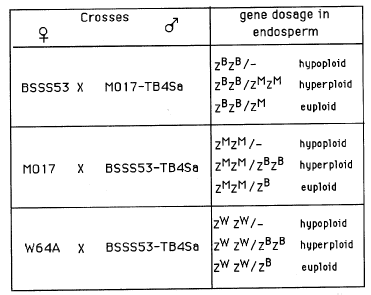
To test whether gene dosage or imprinting of the Mo17 allele accounts for the observed maternal effect, the B-A translocation TB-4Sa, which includes the Zpr10/22 locus, was converted to BSSS53 and Mo17 lines (represented as BSSS53-TB4Sa and Mo17-TB4Sa) and used in various crosses (see Table 1). Fre-quent non-disjunction of the B centromere during pollen gametogenesis causes variations in the dosage of Zpr10/22 contributed by the male parent, leading to recovery of hypoploid, hyperploid or euploid endosperms on the same ear. The hyperploid endosperms arising out of the crosses BSSS53 X Mo17-TB-4Sa and Mo17 X BSSS53-TB-4Sa are identical in Zpr10/22 gene dosage. However, the gametic origin of each allele is different. Comparison of the 10kD zein levels and the 10kD zein mRNA levels between these two groups of hyperploid endosperms is therefore critical in testing whether gene dosage or imprinting accounts for the maternal effect. Since maternal effect is not observed in crosses of BSSS53 and W64A, the cross W64A X BSSS53-TB-4Sa should yield progeny that correlate 10kD zein and its mRNA level with Zpr10/22 dosage.
Our analyses show that the levels of 10kD zein as well as 10kD zein mRNA are similar to BSSS53 in all progeny arising from the cross, BSSS53 X Mo17-TB-4Sa. Likewise, progeny from Mo17 X BSSS53-TB-4Sa are similar to the maternal parent Mo17 in 10kD zein and its mRNA levels. Thus, the expression of 10kD zein appears not to be determined by the dosage of Zpr10/22 in thesecrosses but rather by the gametic origin of the Mo17 allele. However, progeny from W64A X BSSS53-TB4-Sa with varying doses of the BSSS53 allele differ in the levels of 10kD zein expression, suggesting a dosage effect.
Taken together, our results indicate that the observed maternal effect in reciprocal crosses of BSSS53 and Mo17 cannot be explained by considering unequal gene dosage contributions of the parents in the hybrids. This indicates imprinting of the Mo17 allele function whereby it is effective as a dominant downregulator of 10kD zein level only when it is transmitted through the female gametophyte. However, this study cannot determine whether the Mo17 allele is directly imprinted or its function modified in response to other imprinted factors in Mo17. For this, it would be necessary to introgress the Mo17 allele in a non-imprinted line such as BSSS53 or W64A and study the effect of its transmission through the male and female gametophytes.
Figure 1. Western blot analysis of SDS-PAGE fractionated total zein from mature endosperms of BSSS53, W64A and their reciprocal crosses using polyclonal antibody raised against 10kD zein. The arrowhead indicates the 10kD zein position. ZB and ZW represent the Zpr10/22 allele from BSSS53 and W64A respectively.
Figure 2. A. Western blot analysis of SDS-PAGE fractionated total zein from mature endosperms of BSSS53, Mo17 and their reciprocal crosses using polyclonal antibody raised against 10kD zein. The arrowhead indicates the 10kD zein position. ZB and ZM represent the Zpr10/22 allele from BSSS53 and Mo17 respectively. B. Relative levels of 10kD zein mRNA in immature endosperms at various days after pollination (DAP) from BSSS53, Mo17 and their reciprocal crosses. The relative levels were determined by successively probing slot-blot filters of RNA with 10kD zein gene probe and A copy of the 27kD zein gene probe (the levels of 27kD A zein gene mRNA are comparable in BSSS53 and Mo17 at all stages of development). Hybridization intensities obtained with the two probes were determined by densitometry and the level of 10kD zein mRNA normalized against 27kD A gene mRNA. The ratio obtained for Mo17 at DAP 18 was given a value of 1 and all other ratios represented relative to it.
Table
1. Crosses to test gene dosage vs. imprinting of the Mo17 allele. The
B-A translocation TB-4Sa carrying Zpr10/22 on the translocated chromosome
4 arm was backcrossed five times with inbred lines BSSS53 and Mo17 to generate
BSSS53-TB-4Sa and Mo17-TB-4Sa. The expected gene dosage and allelic composition
of Zpr10/22 in the endosperm is indicated next to each cross. ZB,
ZM and ZW represent Zpr10/22 alleles from BSSS53, Mo17 and W64A,
respectively.
Return to the MNL 67 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}