Mutations in the brittle1 (bt1) locus result in a collapsed endosperm phenotype with reduced starch deposition, but the primary defect of mutant bt1 alleles is not known. To provide tools for determining the function of the Bt1-encoded protein, we have isolated molecular clones of the locus using a transposable element-tagged allele (Sullivan et al., Plant Cell 3:1337, 1991). As expected from the endosperm-specific defect of bt1 mutants, Northern blots probed with the cloned Bt1 sequence show that Bt1 transcripts are found in endosperm cells, but not in embryo or leaf tissues.
Analysis of the amino acid sequence, deduced from a wildtype cDNA clone, suggested that the Bt1 protein is plastid localized, since it contains an apparent transit peptide at its amino terminus. Cleavage of the putative transit peptide would produce a 38.6Kd mature protein. Searches of protein sequence data bases indicated that the Bt1 protein has amino acid sequence similarity with several mitochondrial membrane proteins that function in metabolite transport (Sullivan et al., 1991). Taken together these observations suggested that the locus may encode an amyloplast membrane protein involved in metabolite transport.
Experiments to test the function of the transit peptide were performed in Ken Keegstra's laboratory in the Botany Dept. These experiments showed that the Bt1 transit peptide could function in the import of the protein into isolated pea chloroplasts (Li, Sullivan and Keegstra, J. Biol. Chem. 267:18999, 1992). In addition, the imported protein localized to the chloroplast envelope inner membrane. This is consistent with the location of metabolite transport proteins.
To confirm that the Bt1 protein is an amyloplast membrane protein, the subcellular location of the protein was investigated. Antibodies against the carboxy-terminal 56 amino acids of the Bt1-encoded protein were obtained by immunizing a rabbit with a fusion protein produced in E. coli. The fusion protein was produced from a plasmid containing a restriction fragment from the Bt1 cDNA clone inserted downstream of the glutathione-S-transferase gene in the plasmid pGEX-2T (Smith and Johnson, Gene 67:31, 1988). Following purification by affinity chromatography, the anti-Bt1 antibodies react with specific proteins on Western blots of endosperm extracts. These proteins are missing in mutant extracts from the bt1-R allele (data not shown).
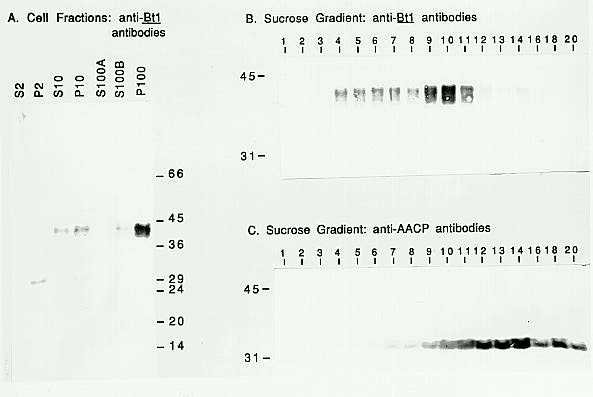
In the summer of 1992, self-pollinated W64A endosperms were harvested (20 days after pollination) and processed to isolate the microsomal membrane fraction using the protocol of Gardner et al. (Physiol. Plant. 69:541, 1987). This fraction should be enriched in amyloplast membranes. In Figure 1A fractions obtained at the different centrifugation steps were probed on a Western blot with the anti-Bt1 antibodies. The microsomal membrane fraction (i.e., the 100,000 xg pellet or P100) is enriched for the protein detected by the anti-Bt1 antibodies. (Actually, Western blots appear to contain more than one Bt1 protein band, with apparent molecular weights ranging from 38 to 42Kd. The basis for this pattern is unknown.)
The P100 was further fractionated by centrifugation through a sucrose density gradient. The gradient fractions were also probed on Western blots (Fig. 1B and C). The Bt1 proteins are found in fractions 4 through 11 in the gradient (Fig. 1B). These fractions also contain the carotenoid pigments of the amyloplast membranes (data not shown). In Figure 1C, a parallel blot of the gradient fractions was probed with antibodies against the mitochondrial membrane ATP/ADP carrier protein (AACP; Hallermayer et al., Eur. J. Biochem. 81:523, 1977). Although there is overlap between the fractions containing Bt1 and AACP, the distribution of the two proteins is clearly different. The migration of Bt1 protein with the carotenoids and in different fractions from a mitochondrial membrane protein suggests that the Bt1 protein is located in the amyloplast membrane.
From these experiments, it seems likely that the Bt1 protein functions as a metabolite transporter. Further experiments will be necessary to determine its metabolite specificity. These results indicate that the deficiency for a starch granule-bound oligosaccharide synthase activity, reported earlier (Pan and Nelson, MNL59:105, 1985), is not the primary lesion of bt1 mutations.
Figure
1. Western blots of endosperm cell fractions. Blots were probed with
the primary antibody as indicated on the panels. Secondary antibody was
phosphatase-linked goat-anti-rabbit IgG, and the blots were stained using
NBT/BCIP. A. Following homogenization, sequential centrifugation of extracts
yielded supernatant (S) and pellet (P) fractions from the 2,000, 10,000
and 100,000 xg steps (2, 10 and 100, respectively). Each lane contains
2mg of protein. B. Sucrose gradient fractions are labeled from the top
of the gradient. An equal volume from each fraction was loaded. C. A parallel
gel was run with the same fractions as used in Panel B.
Return to the MNL 67 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}