Pioneer Hi-Bred International, Inc.
AMES, IOWA
Iowa State University
The influence of random mating on recombination among RFLP loci
--W. D. Beavis, M. Lee, D. Grant, A. R. Hallauer, T. Owens, M. Katt and D. Blair
Studies of recombination in maize have historically used backcross (testcross) or F2 progeny from bi-parental crosses of specially crafted genetic stocks. The development of molecular markers has made it possible to develop saturated linkage maps in adapted germplasm using backcross, F2 and RI progeny. Burr et al. (Genetics 118:519, 1988) have advocated the use of RI populations because there are more meioses to produce detectable recombinants among the progeny. This becomes a desirable goal as the number of genetic loci in a linkage group increases. Another approach to producing a larger percentage of detectable recombinants is to randomly mate the population from a bi-parental cross before obtaining progeny (Kempthorne, ISU Press, 1957). An advantage of random mating is that the frequency of detectable recombinants will ultimately approach 0.5 regardless of how tightly linked the loci initially appear. It is also possible to keep linkage at some level less than 0.5 by simply discontinuing random mating at some intermediate generation and deriving either F2 or RI progeny.
We have developed linkage maps using RFLP-loci and F2 progeny from randomly mated and non-randomly mated populations. Both populations were derived from a cross of B73xMO17. The non-randomly mated population, designated syn0, was obtained by selfing the original F1 and consists of 99 progeny. These progeny were further selfed to produce syn0-F2:3 lines. The randomly mated population, designated syn5, was derived by sib-mating the original F1 for five cycles, then deriving 98 syn5-F2:3 lines as in the syn0 population. RFLP-typing of these lines was accomplished using standard molecular biology protocols. The frequency of the RFLP alleles and segregation at the loci were analyzed for deviations from expected frequencies based on Mendelian inheritance. The genetic linkage maps were constructed using MAPMAKER (Lander, E et al., Genomics 1:174, 1987) and a mapping protocol similar to that outlined by Landry et al. (Genome 82:543, 1991).
There was no evidence for segregation distortion at any of the loci in the syn0, but there was evidence of distorted segregation at 5 loci in the syn5 (Table 1). The frequency of the B73 allele at most (24) of the loci in the syn0 was less than 0.5, but only umc76 was significantly less. Interestingly, the frequency of the B73 allele at most (23) of the loci in the syn5 was greater than 0.5. The frequency of the B73 allele at five loci was greater than 0.60 and at two loci it was less than 0.40. The change in frequency from the syn0 generation to the syn5 generation was almost always toward the B73 allele. However, for bnl8.10, pio200518, bnl7.25, and bnl8.29, the change was toward the MO17 allele. These loci were all linked in the syn0 generation (Figure 1). Interestingly, a plant height QTL was identified in this region using data from the syn0 population (data not shown).
Table 1. Results of segregation analyses (Chi-squared statistic) and
frequencies of the B73 allele for 32 RFLP-loci in F2 progeny derived from
randomly mated (syn5) and nonrandomly mated (syn0) populations of B73xMO17.
| Calculated Chi-squared values | Frequency of B73 allele | |||
| LOCUS | syn0 | syn5 | syn0 | syn5 |
| umc94 | 4.12 | 5.06 | 0.44 | 0.58 |
| bnl5.62 | 4.52 | 2.56 | 0.42 | 0.56 |
| pio200537 | 4.90 | 7.04 | 0.46 | 0.43 |
| pio200603 | 11.80 | 6.08 | 0.43 | 0.44 |
| pio200689 | 2.98 | 0.92 | 0.47 | 0.47 |
| pio200640 | 6.20 | 7.85 | 0.42 | 0.54 |
| umc76 | 9.50 | 1.04 | 0.38 | 0.52 |
| umc137 | 2.17 | 1.41 | 0.45 | 0.53 |
| umc11 | 3.38 | 2.21 | 0.44 | 0.53 |
| umc13 | 2.90 | 0.56 | 0.44 | 0.53 |
| umc1C12 | 1.73 | 24.04 | 0.45 | 0.66 |
| bnl7.21 | 2.25 | 14.64 | 0.45 | 0.62 |
| umc133 | 2.90 | 8.71 | 0.44 | 0.57 |
| pio200654 | 3.73 | 16.51 | 0.50 | 0.63 |
| bnl5.59 | 5.02 | 8.86 | 0.43 | 0.61 |
| pio200575 | 4.02 | 1.12 | 0.43 | 0.53 |
| pio200674 | 3.04 | 2.54 | 0.44 | 0.55 |
| pio200644 | 2.60 | 3.31 | 0.46 | 0.50 |
| pio200855 | 3.71 | 0.40 | 0.45 | 0.49 |
| umc23 | 5.04 | 4.39 | 0.51 | 0.54 |
| umc33 | 3.72 | 0.25 | 0.45 | 0.51 |
| pio200668 | 3.30 | 0.06 | 0.45 | 0.51 |
| pio200661 | 1.50 | 1.05 | 0.47 | 0.51 |
| umc128 | 1.43 | 3.20 | 0.46 | 0.49 |
| umc50 | 1.38 | 0.10 | 0.53 | 0.49 |
| bnl8.10 | 0.34 | 30.75 | 0.51 | 0.36 |
| pio200518 | 2.60 | 0.91 | 0.52 | 0.47 |
| bnl7.25 | 1.88 | 4.74 | 0.52 | 0.43 |
| bnl8.29 | 6.50 | 10.75 | 0.52 | 0.39 |
| umc84 | 0.38 | 18.61 | 0.51 | 0.64 |
| pio200557 | 3.32 | 10.68 | 0.48 | 0.62 |
| bnl6.32 | 4.46 | 8.05 | 0.48 | 0.58 |
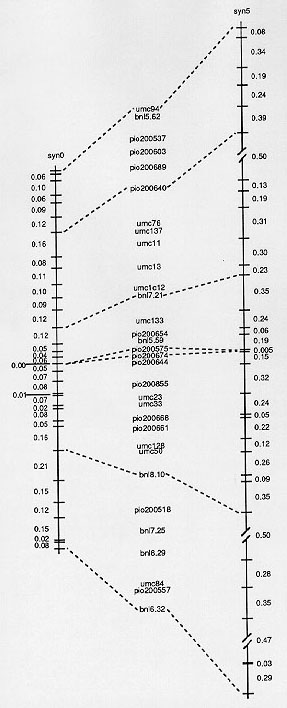
Figure 1. RFLP linkage maps of chromosome 1 based on segregation data from F2 progeny derived from an F1, (syn0) and an F1 randomly mated for five generations (syn5) of the cross B73xMO17.
The sum of the estimated recombination among adjacent pairs of RFLP loci in the syn0 population was ~ 275. This value is unadjusted for expected double recombinants. Thus, the estimated average recombination was ~ 8.8. A comparison of the linkage maps (Figure 1) revealed that there are four independent linkage groups after 5 cycles of mating. The sum of the estimated recombination among adjacent pairs of RFLP loci in the syn5 population was ~ 730 (unadjusted for expected double recombinants), or an average of about 23.5. The relationship of recombination among randomly mated generations can be described
r(n) = r(n-1) + [r(n-1) - 2r2(n-1) ]/2, (1)
where r(n) is the frequency of recombinants after n generations of random mating and r(n-1) is the frequency of recombinants in the prior generation. Based on the recurrence relationship, (1), and the estimated recombination in syn0, the average frequency of recombinants should have been about 35 in syn5.
The estimated recombination in the syn5 was greater than the syn0 for all pairs of adjacent loci. For many pairs of adjacent loci the observed value for r(5) was about what would be expected based upon the estimate from syn0 and (1). For example, the estimated recombination between pio200661 and pio200668 was 1.5% in the syn0. Assuming that this is the true value for this segment, then the expected amount of recombination in the syn5 would be ~10%; we observed 12%. On the other hand, there were several segments where the observed recombination in the syn5 was much less (10-20%) than expected based on the observed values in the syn0 (Figure 1). For example, there was very little increase in the number of recombinants between umc94 and bnl5.62.
One of the primary objectives for generating a randomly mated population
is to uncouple tight linkages. In the syn0, we were unable to detect recombinants
between pio200575 and pio200674. In two additional B73xMO17
populations, based on data from 112 F2's and 46 RI's, we have not been
able to detect recombinants between these loci (data not shown), suggesting
very tight linkage. However, in the syn5, we did detect one recombinant
between these loci among the 196 sampled gametes. Uncoupling tight linkages
also will be useful for QTL studies. Beavis, WD et al. (1989 presentation
to ASA, Las Vegas, NV) used simulations to show that randomly mated populations
improve resolution of QTL. Patterson, EB et al. (Genetics 124:735, 1990)
advocated random mating of isogenic lines as a basis for map-based cloning
of introgressed genetic loci. Finally, to be politically correct, we note
that the ease with which random mating is accomplished in maize makes it
an ideal organism for these types of recombination studies.
Return to the MNL 66 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}