Indiana University
Two ring chromosome derivatives containing a duplication of the R locus
--Ellen Dempsey and M. M. Rhoades
The ring-10 chromosome of Emmerling was produced following irradiation of a K10-I chromosome when breaks occurred near the ends of 10S and 10L (Emmerling, Genetics 40, 1955). A series of ring-10 derivatives was studied by Miles (Indiana University Ph.D. Thesis, 1970). Among these was the Km chromosome in which a long duplication, including the R and G loci as well as the three prominent chromomeres and a portion of the large knob of K10-I, was attached to the short arm of chromosome 10. In this configuration, one attribute of K10-I was lost, namely the induction of preferential segregation, although the crossover enhancement effect of K10-I was still operating. Derivatives of Km in which the knob and three chromomeres were restored to their original orientation in 10L recovered some or most of the preferential segregation activity.
Two new derivatives arose in our stocks of ring-10; both resemble Km in having a duplication of the R locus in 10S. One has the truncated knob of K10-I and the three chromomeres and was designated Km-medium since it is shorter than Km. The other has the same knob plus a small euchromatic tip terminating the short arm but lacks the three chromomeres; it was called Km-short. Both chromosomes have a normal knobless 10L.
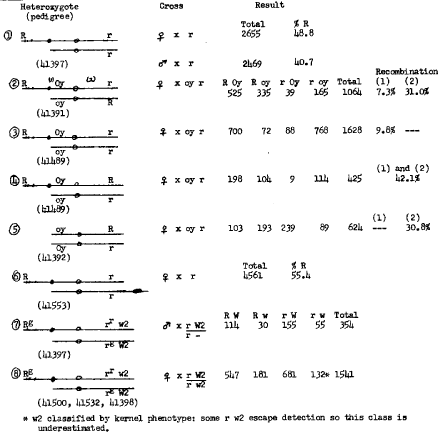
Data from genetic studies with Km-medium are presented in Table 1. Transmission of the duplicated chromosome marked by R was followed in testcrosses (item 1 of Table 1). Female transmission was very slightly reduced and male transmission was about 40%. The longer Km chromosome of Emmerling and Miles gave 40% female transmission and only 6-7% male transmission. The R locus in the duplication shows close linkage with oy in 10S (crosses 2 and 3). The 7-10% recombination observed was much less than the control value of 30.8% in plants without the duplication. Compounds having R Oy r/oy r, R Oy R/oy r, and R Oy r/oy R constitution were consistent in giving recombination of approximately 10% for the duplicate R-oy segment and about 30% for the oy-r region (crosses 2, 3, and 4). When Km-medium was opposed by a K10-I carrying the r allele, female transmission of R on the duplication chromosome was considerably increased (cross 6). This result was unexpected since female testcrosses of K10-I r/N10 R heterozygotes give 70% r due to preferential segregation. However, the increased transmission of the duplicated chromosome in Km-medium R r/K10-I r compounds becomes understandable when the output of single crossovers in 10S and 10L is considered. Singles in 10S give heteromorphic dyads at Anaphase I with the chromatids containing the duplicated R oriented toward the poles. Preferential recovery of the duplicated chromatid in the basal megaspore is expected. Single crossovers in 10L between the centromere and K10 knob would normally result in preferential segregation for the K10-I chromatid marked by r, but half of such chromatids also possess the duplicated R by virtue of the crossover so the ratio of R:r following singles in 10L remains 1:1.
Table 1. Crosses of Km-medium heterozygotes.
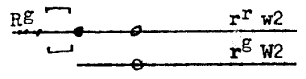
Surprising results were observed in cross 7 of Table 1. Previous studies (Rhoades and Dempsey, Plant Genetics, UCLA Symposium, 1985) have shown that the K10-I chromosome has a segment from the distal region of 10L inserted in inverted order between the three-chromomere region and the large knob of K10-I. Three loci, L13, O7 and W2, are included in this segment. In cross 7, an r tester hemizygous for W2 was pollinated by a plant heterozygous for the Km-medium chromosome with the duplicated R. The progeny included kernels displaying the typical w2 phenotype (colored kernels with multiple depressed colorless sectors and colorless kernels with scarred floury-like sectors). When planted, these kernels gave white seedlings. The male parent apparently had the genotype (Figure 1)
The w2 plants derived from R kernels were unexpected since R should be almost completely linked with W2 in the bracketed region. We conclude that either (1) the W2 locus is not present in the duplication or (2) the W2 locus is inactive in the new location. The R allele in the duplication retains its full activity in the seed component. The original ring and the Km of Miles and Emmerling had the R-r allele (with a dominant P component). Our crosses suggest that the duplication in Km-medium contains R-g; possibly activity of the P component is also suppressed in the new location. We hope to discriminate between alternatives (1) and (2) by transferring the duplicated segment back to 10L and noting whether the W2 and P component of R regain their activity. To achieve this transfer, a rare crossover must occur which involves pairing of the duplication with a modified 10L containing the three-chromomere region of K10-I but lacking W2 (Df(F)). The unusual results of cross 7 have been reinforced by the finding of scarred variegated R kernels (called R-spotted) in the cross of Km-medium heterozygotes by r W2/r w2 pollen parents (cross 8). The proportion of scarred kernels (both R and r) on an ear is close to 25%, with roughly equal numbers in the colored and colorless classes. Again, there is no evidence of a W2 allele in the duplicated segment.
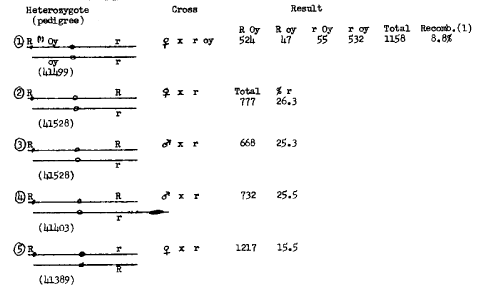
Table 2 shows the data derived from crosses of Km-short heterozygotes. Cross 1 illustrates the close linkage of R and oy, similar to that found with Km-medium. The percentage of R indicates normal female transmission of the duplicated chromosome. The duplicated region is considerably shorter than in Km-medium and the R locus has been transposed to a position close to the knob. Crosses 2 and 3 show the results of male and female testcrosses when the duplicated chromosome carries two R alleles. The r kernels come only from noncrossovers and from crossovers distal to R in the long arm. A similar percentage of r kernels was obtained in cross 4 where the duplicated R R chromosome was opposed by K10-I carrying r. No preferential segregation is expected in the male testcross. In cross 5 there is a hidden r allele which is revealed in the progeny following a crossover anywhere between the R and r alleles on the duplicated chromosome. Thus, the r kernels represent one half of such crossovers and the chromosome bearing two R alleles (the reciprocal crossover) is not detected. Results in Table 1 with Km-medium indicated a map distance of about 40 between the two R loci. If similar recombination frequencies occur in Km-short heterozygotes, one would expect 20% r (crossovers) in cross 5 and 30% r (noncrossovers and distal crossovers) in crosses 2, 3, and 4. The lower frequencies observed for both crossover and noncrossover classes are difficult to explain.
Finally, both chromosomes were tested for their ability to induce preferential segregation in chromosome 9. The data are not extensive (see Table 3) but no preferential segregation of the yg2 allele in chromosome 9 was observed. No change was noted due to the presence or absence of a Df(K) chromosome which carries the differential segment (three-chromomere region) but not the knob of K10-I. A frequency of 65% yg has been observed in similar heterozygotes carrying an intact K10-I chromosome. The lack of preferential segregation observed with the two ring chromosome derivatives is in agreement with Miles's observation that the original Km does not induce preferential segregation. Thus, transfer of the knob to the short arm of chromosome 10 eliminates its capacity to induce preferential segregation in other chromosome pairs and may also affect the activity of genes in the duplicated segment.
Table 2. Crosses of Km-short heterozygotes.
Table 3. Tests for preferential segregation.
Return to the MNL 66 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}