--Nina Fedoroff, Michael Schläppi, Douglas Cook and Vladimir Kolosha
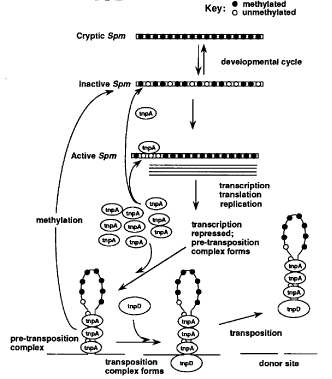
The accompanying diagram illustrates our present view of the manner in which Spm transcription and translation are regulated. In previous reports from this laboratory, we showed that inactive Spm elements can exist in an extremely stably inactive form which we designated cryptic and a much less stably inactive form that can be distinguished genetically by its ability to be reactivated upon introduction of an active Spm element (Fedoroff, NV and Banks, JA, Genetics 120:559, 1988; Banks et al.,Genes Dev. 2:1364, 1988; Fedoroff, Genetics 121:591, 1989). Cryptic elements are more extensively methylated within the GC-rich region of the first exon, termed the downstream control region (DCR), than are less stably inactive elements. The methylation levels of both cryptic and inactive elements are lower in embryonic DNA than in DNA of seedlings or mature plants, suggesting that there is a reduction in methylation early in development (Banks and Fedoroff, Dev. Genet. 10:425, 1989). Inactive Spm elements exhibit different methylation levels in different plant parts during development and methylation of both cryptic and inactive elements is reduced by the introduction of an active element (Banks, JA et al., 1988; Banks and Fedoroff, 1989). We have shown here and elsewhere that genetically inactive elements are transcriptionally silent, but can be transcribed in the presence of an active element (Banks et al., 1988; Kolosha, V and Fedoroff, this volume). The accompanying reports by Cook, D and Fedoroff and by Schläppi, M and Fedoroff present evidence suggesting that the Spm promoter is inhibited by methylation, as well as by high levels of TnpA in both a transient assay system and in stably transformed tobacco cells. However, introduction of a tnpA cDNA into tobacco cells containing a resident Spm element stimulates excision of a dSpm element.
As shown in the accompanying diagram, these observations lead us to propose that the TnpA protein exerts two kinds of regulatory effect, one which promotes and one which represses element transcription. At low concentrations, we hypothesize that TnpA stimulates element expression. Whether it acts directly as a transcription factor or indirectly by preventing methylation has not yet been determined, although we have suggested that it may do both (Masson, P et al., Genetics 177:117, 1987; Banks, JA et al., 1988). Nor do we know whether TnpA acts directly by binding to methylatable sites or indirectly by maintaining the element in a conformation unfavorable for methylation. Both types of interference may play a role, since the tnpA protein's recognition sequences are confined to the element's subterminal repetitive regions and contain methylatable C residues (Masson et al., 1987; Gierl, A et al., EMBO J. 7:4045, 1988), yet changes in methylation of the adjacent GC-rich DCR are also observed in reactivating elements. Gierl et al. (1988) have reported that methylation of C residues within the TnpA binding sequence reduces the affinity of the binding site for the protein, which may be an important factor in regulating element expression.
Thus it appears that element methylation is low early in development (as illustrated in the top part of the diagram), perhaps by virtue of a general suppression of DNA methylation. Remethylation and TnpA binding may both occur early in development, but the presence of TnpA protein clearly favors reduced methylation and element expression, whatever the molecular mechanism. As the concentration of TnpA increases, it may occupy progressively more of the binding sites, eventually reaching a level at which further transcription is repressed and the element ends are brought together in a complex, which we have designated the "pre-transposition complex" in the accompanying diagram (Frey et al., EMBO J., 12:4037, 1990; Masson, P et al., Plant Cell 3:73, 1991). The present model elaborates on previous models (Masson et al., 1987; Banks, JA et al., 1988; Gierl et al., 1988; Frey et al., 1990; Masson et al., 1991) in postulating that the pre-transposition complex has one of two alternative fates. The first is to bind the TnpD protein, which has been shown to be required for transposition together with the TnpA protein (Frey et al., 1990; Masson et al., 1991), forming a transposition complex that leads to excision of the element from the donor site. The second is to to be remethylated and return to an inactive form. Element inactivation late in development can be monitored by the appearance of sectors on kernels of the appropriate genetic constitution (containing, for example, an Spm-suppressible allele and a trans-acting Spm) and occurs at a high frequency (Masson et al., 1987; also unpublished observations).
We propose that the rate-limiting step in the transposition process is the formation of the pre-transposition complex for the several reasons that follow. First, we have observed that the introduction of a tnpA cDNA into tobacco cells with an Spm element increases the excision frequency, but introduction of a tnpD cDNA does not (Schläppi and Fedoroff, this volume). This implies that it is the TnpA protein that is rate limiting in these cells. Moreover, deletions that encroach on the subterminal repeats of the Spm element reduce the transposition frequency and delay it in development (Schwarz-Sommer et al.,EMBO J. 10:2439, 1985), suggesting that an element missing some of the TnpA binding sites is less efficient in forming the pre-transposition complex, perhaps requiring higher TnpA concentrations than elements with the full complement of subterminal repeats. The third and perhaps most compelling argument is a genetic one, based on the phenotypes of Spm-suppressible alleles. The predominant phenotype of plants containing both an Spm and such an Spm-suppressible dSpm insertion mutation is that characteristic of the dSpm mutant in the presence of an active element. This suggests that the active Spm element generally remains in a transcriptionally active form, because when the trans-acting Spm element is lost (by virtue of chromosome breakage, for example), the phenotype of the monitoring dSpm insertion mutation immediately exhibits the phenotype that is characteristic of it in the absence of a trans-acting element. This implies that the concentration of TnpA drops rapidly when the element is removed or inactivated and that complexes that form between TnpA and the element are not stable. Thus transcriptional inactivation of the element should be perceptible as sectors of the "no Spm" phenotype. As noted above, such sectors are frequent, although generally small for a fully active Spm, implying that inactivation is a late event in development. Moreover, they never include revertant wildtype sectors within their perimeter, which implies that dSpm transposition does not occur when the element is not expressed. Thus we suggest that transcriptional inhibition must either be followed immediately by transposition or rapidly result in complete inactivation of the element by virtue of the instability of the complex or the TnpA protein (or both). This, in turn, would release the element in a methylatable, TnpA-free form.
An observation made by one of us is perhaps relevant in this context (NF, unpublished). In a study conducted some time ago, it was noted that a majority of kernels containing an active Spm element and a monitoring dSpm mutation and selected for evidence of Spm inactivation gave plants with a transposed (inactive or reactivating) element, rather than an inactive Spm at its original location (these occur as well, but very infrequently). Although it cannot be ruled out that the reinsertion site is responsible for inactivation of the element, it is possible that the more frequent inactivation of newly transposed elements reflects the transcriptionally inactive structure of the transposition complex postulated here. A satisfying aspect of the present model is that it provides an explanation for the origin of the transcriptionally inactive, methylatable state as a by-product of the transposition process. Thus we perceive the transcriptional silencing of the element and the subsequent stabilization of the silent state by methylation as separate events. Moreover, it provides an explanation for the low frequency with which an active element undergoes germinal inactivation.
Note: the authors welcome your comments on this model.
Return to the MNL 66 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}