Carnegie Institution of Washington
Transcriptional reactivation of inactive Spm elements in the presence of Spm-w elements
--Vladimir Kolosha and Nina Fedoroff
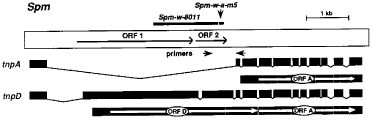
The 8.3kb maize suppressor-mutator (Spm) transposable element comprises a single transcription unit and its primary transcript gives rise to several processed, polyadenylated mRNAs by alternative splicing (Masson, P et al., Genetics 177:117, 1987; Masson et al., Cell 58:755, 1989). The mRNAs potentially encode at least 4 different polypeptides, some with extensive common domains (Masson, P et al., 1989). Two element-encoded proteins, designated TnpA and TnpD, have been shown to be required for transposition of the element (Frey et al., EMBO J. 9:4037, 1990; Masson et al., Pl. Cell 3:73, 1991). The structure of the tnpA and tnpD transcripts is shown in Figure 1, along with the structure of the element. The 68kDa DNA-binding TnpA protein is encoded by the most abundant transcript of the element (Pereira et al., EMBO J. 5:835, 1986; Gierl, A et al., EMBO J. 7:4045, 1988). The TnpD protein is encoded entirely within the first intron of the tnpA transcript, which contains two long open reading frames (ORF1 and ORF2) and is expressed from a rare, alternatively spliced dicistronic transcript which also codes for TnpA (Masson, P, P et al., 1989). Spm elements are subject to both positive and negative regulation. Spm can be inactivated by a heritable, reversible genetic mechanism (Fedoroff, NV and Banks, JA, Genetics 120:559, 1988). Inactivation is correlated with increased methylation of C residues both upstream and downstream of the element's transcription start site. McClintock, B reported that an inactive Spm (Spm-i) element is subject to transient genetic reactivation in the presence of an active element, suggesting the existence of a positive autoregulatory gene on the element (McClintock, CIW Yrbk 57:415, 1957). Two Spm-w elements with deletions eliminating part of the TnpD coding sequence have been shown to retain the ability to trans-activate an inactive element genetically (Fedoroff, NV and Banks, JA, 1988; Fedoroff, Genetics 121:591, 1989).
We showed that tnpA transcripts are more abundant in plants containing both an inactive element and a trans-activating Spm-w element than in plants with only one of the two elements (Banks, JA et al., Genes Dev. 2:1364, 1988). In these experiments, carried out before the element's minor, alternatively spliced transcripts had been identified and characterized (Masson, P et al., 1989), transcripts of the trans-activating element were not distinguished from those of the activated element. By making use of inactive and trans-activating elements which can be distinguished structurally, we show here that an Spm-i is transcribed in the presence of an active one. We have used two different Spm-w elements, both of which have internal deletions, as the trans-activating element. The structure of the standard Spm (Spm-s) and the Spm-w elements is shown in Figure 1. The Spm-w-8011 element has a 1.7kb internal deletion commencing in ORF1 and ending in ORF2, while the Spm-w-a-m5 element has a 144bp deletion near the 3'end of ORF2 (Masson, P et al., 1987; Masson et al., 1989). The inactive element is an Spm-s element inserted at a site just upstream of the a gene (Masson, P et al., 1987). The element has undergone spontaneous genetic inactivation, which we have previously shown to be associated with an increase in the extent of methylation of C residues in the immediate vicinity of the element's transcription start site (Fedoroff, NV and Banks, JA, 1988; Banks et al., 1988; Banks and Fedoroff, Dev. Genet. 10:425, 1989).
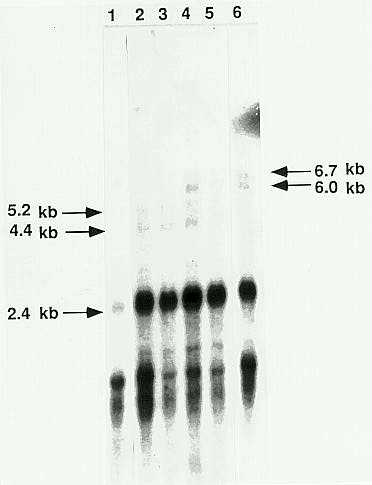
Several size classes of transcripts with homology to the Spm element can be detected in maize plants with active elements (Masson, P et al., 1989). The most abundant transcript encoded by the intact standard Spm element is the approximately 2.4kb tnpA transcript (Figure 2, lane 6). Plants with active elements also have larger and smaller transcripts that show homology to the element. Two size classes of large transcripts have been detected (Masson et al., 1989). From careful measurements of relative mobility, the sizes of these transcripts are presently estimated to be 6.0 and 6.7kb. The shorter of these transcripts therefore corresponds in length to the previously identified tnpC and tnpD transcripts, while the longer exceeds this length and may represent a partially processed transcript. Short transcripts with homology to the Spm element are also abundant in maize plants. Many short transcripts have been cloned and found to be processed transcripts of internally deleted, transposition-defective elements. Transcripts with the length expected for the previously described tnpB transcript are not detectable by blot hybridization (Masson, P et al., 1989).
None of the large-element-encoded transcripts can be detected by blot hybridization in RNA isolated from plants with an Spm-i element (Figure 2, lane 1), but a small amount of tnpA transcript can be detected in many such plants. Short Spm-homologous transcripts are as abundant in plants with an Spm-i as in those with an active element. In RNA isolated from plants containing the Spm-w-8011 element as the only genetically active element, the two largest detectable Spm-homologous transcripts have estimated lengths of 5.2 and 4.4kb (Figure 2, lanes 2 and 3). These size estimates are in reasonably good agreement with the anticipated length of the large transcripts from the deleted element. The length of the tnpA transcript is indistinguishable between RNA preparations from plants with the Spm-w-8011 and Spm-s elements. This suggests that processing of transcripts of the Spm-w-8011 element is not affected by the deletion. The large transcripts from both the Spm-s and the Spm-w-8011 elements are detectable in RNA from plants that contain both the Spm-w-8011 element and an Spm-i element (Figure 2, lanes 4 and 5). The relative intensities of the bands corresponding to the transcripts of the deleted Spm-w-8011 element and the Spm-s elements are approximately the same, suggesting that the large transcripts of the two elements are roughly comparable in abundance.
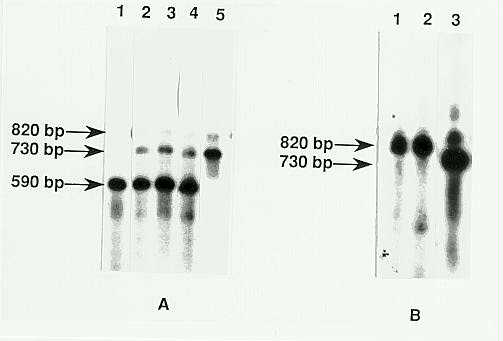
Similar results have been obtained with RNA from plants containing the Spm-w-a-m5 element and an Spm-i element, using PCR amplification of internal cDNA fragments to distinguish between the trans-activating deleted element and the resident, standard Spm-i element. Primers flanked the deletion in the Spm-w-a-m5 element in order to detect tnpD transcripts from the deleted and standard elements. These give rise to a 730bp DNA fragment from the completely processed tnpD transcript of the standard element and a 590bp fragment from the deleted Spm-w-a-m5 element (Figure 3A, lanes 1 and 5). Direct sequencing of the PCR-amplified fragments confirmed that they were derived from the completely processed tnpD transcript of the standard and deleted elements. Variable amounts of a longer, Spm-homologous 820bp fragment have been detected (Figure 3). The fragment was sequenced and determined to contain the third intron of the tnpD transcript. Because incorporation of DNAse into the reaction mixture prior to cDNA synthesis eliminated this band, we conclude that it is attributable to DNA contamination.
Using the extremely sensitive PCR amplification reaction, we have detected only the 590bp fragment expected for the tnpD transcript of the Spm-w-a-m5 element in plants containing this element as the only genetically active Spm (Figure 3A, lane 1). Moreover, the 730bp fragment expected from transcripts of the standard Spm element was detected in only 1 of seven different plants containing an Spm-i element. In RNA isolated from 6 of the 7 plants, the only fragment that could be amplified was the 820bp fragment derived from contaminating genomic DNA (Figure 3B, lanes 1 and 2). The fragment expected for the tnpD transcript of the standard Spm element was detected in the seventh plant with two different primer pairs. Since the plants used in the present experiments contained an unstably inactive element, it is likely that the element in this plant had undergone spontaneous reactivation. By contrast, plants containing both an inactive Spm-s and the Spm-w-a-m5 invariably gave cDNA preparations from which both the 730bp and 590bp fragments could be amplified (Figure 3, lanes 2-4). This implies that tnpD transcripts of both elements are present in such plants. The 590bp fragment characteristic of the tnpD transcript of the Spm-w-a-m5 element was generally more abundant in such amplification reactions than the 730bp fragment expected from the Spm-s tnpD transcript. But because the abundance of a given fragment among the final amplification products is influenced by its length, we cannot conclude that the transcript of the deleted element is more abundant than that of the intact element.
To obtain a rough estimate of the extent to which the inactive element had been reactivated in plants containing an Spm-w element, we quantified the amount of tnpA transcript relative to the amount of actin transcript by scanning Northern blots (Shah et al., J. Mol. Appl. Genet. 2:111, 1983). The abundance of tnpA transcripts in six plants with a single Spm-i element ranged from 2% to 30% (average of 12±10%) of its abundance in a plant containing a single Spm-s element. Three plants with an Spm-w-8011 element contained an average of 35±20% as much tnpA transcript as plants with a standard element. The abundance of the tnpA transcript in RNA from plants containing a single copy of the Spm-w-a-m5 element is 70±35% of its abundance in RNA from plants with a single Spm-s element. Eight pairwise comparisons were done between sibling plants containing only the Spm-w-8011 element and both the Spm-w-8011 element and an Spm-i element. The plants with both elements contained 2.8 (±0.8) times as much tnpA transcript as plants containing only a weak element. Similar pairwise comparisons between mRNA preparations from eight pairs of plants containing either only the Spm-w-a-m5 element or both the Spm-i element and an inactive element yielded a tnpA transcript ratio of 1.8 (±0.5). Although the plant-to-plant variation in the abundance of element-specific transcripts is considerable, the results of these experiments suggest that the tnpA transcript is about as abundant in plants with an Spm-w-a-m5 element and somewhat less abundant in plants with an Spm-w-8011 element than in plants with a standard element. Whether this disparity is attributable to a difference in element transcription or in the stability of the transcript is not known. Since neither deletion alters the structure of the tnpA transcript, the effect on the abundance of the transcript may be indirect. Plants with an unstably inactive element contain low, but detectable amounts of tnpA transcript. The 2-3-fold greater amount of tnpA transcript in plants with both an inactive Spm element and a trans-activating Spm-w element suggests that the inactive element is fully active in the presence of the Spm-w element.
The results of present experiments provide direct evidence that transcripts
of an Spm-i element are more abundant in plants that also contain
an active element. We conclude that the capacity of an active Spm element
to reactivate an inactive one, first inferred from genetic experiments,
is detectable at the transcriptional level. We have previously reported
that unstably inactive elements are more extensively methylated than active
elements and we have noted that the extent of methylation is markedly lower
in plants containing a trans-activating Spm-w element (Banks, JA
et al., 1988). These results imply an interaction between a gene product
encoded by the Spm-w element and the inactive element. They do not
permit us to distinguish between a direct transcription factor-like effect
and an indirect effect mediated by a reduction in methylation.
Figure 1. A diagrammatic representation of the Spm transposable element. The exon structure and ORFs of the two element-encoded transcripts for which functions have been established, tnpA and tnpD, are represented below the diagram. Located within the first intron of the tnpA transcript are two large open reading frames (ORF1 and ORF2) of which ORFD, the coding sequence for TnpD, is comprised. ORFA, the TnpA coding sequence, is encoded by the last 10 exons and is encoded in both the tnpA and tnpD transcripts. Bars just above the diagram show the location and extent of the deletions in the Spm-w-8011 and Spm-w-a-m5 elements. Arrows below the diagram show the location of the PCR primers.
Figure 2. Northern blot hybridization analysis of polyA+ RNA isolated from plants containing an inactive element (lane 1), the Spm-w-8011 element (lanes 2 and 3), both an inactive element and the Spm-w-8011 element (lanes 4 and 5), and an Spm-s (lane 6). The blot was probed with a cloned, full-length Spm DNA fragment.
Figure
3. Southern blot hybridization analysis of fragments amplified by PCR
from cDNA prepared by reverse transcription of polyA+ mRNAs. A. The RNAs
were isolated from plants with an Spm-w-a-m5 element (lane 1), both
an Spm-w-a-m5 element and an Spm-i element (lanes 2, 3 and
4) and an Spm-s element (lane 5). B. RNA was isolated from plants
with an inactive Spm (lanes 1 and 2) or an active Spm (lane
3). The blot was probed with a DNA fragment homologous to ORF2.
Return to the MNL 66 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}