--Martha G. James, Michael J. Scanlon, Minmin Qin, Donald S. Robertson and Alan M. Myers
This report characterizes Mu-A2, a transposon of the Mutator family known to provide transposition function to an independent Mu element, Mu1, located within the a1-mum2 allele of the A1 gene. A genetic element controlling Mu1 transposition was identified in specific maize lines in which mutable and stable a1 kernels were produced at a 1:1 ratio in crosses to standard lines (Robertson, DS and Stinard, PS, Dev. Genet. 10:482-506, 1989). This genetic element is termed here Mu-A2. Subsequent genetic analysis showed Mu-A2 is a mobile locus and, furthermore, that its copy number can increase (Robertson and Stinard, Theor. Appl. Genet. 82: in press). These results suggested the Mu-A2 locus represents a transposon capable of regulating Mu1. Other studies also identified genetic loci controlling a1-mum2 mutability; these elements, termed Cy (Schnable and Peterson, Mol. Gen. Genet. 215:317-321, 1989), and Mu-R1 (Chomet et al., Genetics 129:261-270, 1991) may be the same genetic element as Mu-A2.
The same strain used to define Mu-A2 genetically was also used to identify and isolate a specific genomic DNA fragment that cosegregated with a1-mum2 mutability (Qin and Ellingboe, Mol. Gen. Genet. 224:357-363, 1990; Qin et al., Genetics 129:845-854, 1991). An increase in the copy number of this genomic fragment correlated with an increased degree of mutability. Taken together, these data indicated the cloned fragment contains the Mu-A2 locus. Nucleotide sequence analysis identified a Mu element within this fragment based on its high degree of homology to previously described Mu elements in the regions of the terminal inverted repeats (Qin et al., Genetics 129:845-854, 1991). The complete 4942 base pair (bp) nucleotide sequence of Mu-A2 is contained in the EMBL sequence database (accession number X62251).
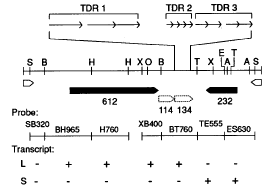
Sequence analysis of Mu-A2 revealed two ATG-initiated open reading frames (ORF) that could encode a polypeptide of more than 100 amino acids in length. The largest of these is a 612 codon ORF (ORF612, from nt 970 to nt 2805) known from Northern hybridization analysis to be transcribed into a 3.4 kilobase (kb) mRNA, denoted transcript "L" (Qin and Ellingboe, Mol. Gen. Genet. 224:357-363, 1991; Figs. 1 and 2). This transcript is present specifically in plants derived from mutable kernels in a cross of Mu-A2 and wildtype strains (Qin and Ellingboe, Mol. Gen. Genet. 224:357-363, 1990; this report). Two additional ORFs (ORF115, from nt 2806 to nt 3145, and ORF134, from nt 3146 to nt 3547) are located immediately downstream of ORF612 and may be present on the same 3.4kb transcript (Fig. 2). Neither ORF115 nor ORF134 contains an initiation codon; thus, their expression as protein would depend on RNA splicing or a translational mechanism that attached this coding information to ORF612. Also, RNA splicing in the region upstream of ORF612 may enable transcription to initiate from an ATG codon located 5' to this ORF, thus extending the reading frame. A second ATG-initiated ORF (ORF232, from nt 4477 to nt 3782) is transcribed on the opposite strand from ORF612 into a 900bp mRNA, denoted transcript "S" (Qin and Ellingboe, Mol. Gen. Genet. 224:357-363, 1990; Fig. 1). Neither transcript "L" nor transcript "S" specifies a polypeptide homologous to any sequence in the Protein Identification Resource database. An interesting feature of ORF134 is the clustering of three different tandem direct repeat (TDR) sequences comprised of 55nt (TDR1), 11nt (TDR2), and 27nt (TDR3) (Fig. 1). These repeats are situated so that the 3'nt of ORF115 is the 5' end of the first repeated sequence, TDR1 (Fig.1). The repeated sequences are not similar to direct repeats found in Mu1, Mu1.7, or rcy:Mu7.
Figure 1. The 4942nt sequence of Mu-A2 is represented by a solid line with restriction sites indicated for SacI (S), BamHI (B), XbaI (X), XhoI (O), StuI (T), EcoRI (E), and SalI (A). Open polygons show terminal inverted repeats, solid polygons show ORFs beginning with ATG initiation codons, and dotted polygons show ORFs lacking an ATG codon. Numbers under each ORF indicate codon length. Polygons point in the 5' to 3' direction of the element they represent. The expanded area comprises 338nt; arrows represent tandem direct repeats. All elements are drawn to scale. Hybridization probes in Northern analyses were random-primer labeled and are identified by appropriate restriction enzyme letter codes followed by numbers indicating fragment nucleotide length. Hybridization of a particular probe fragment to either transcript "L" or transcript "S" (see text) is denoted as "+" or "-", respectively.
To date, Mu-A2, Mu-R1, and Cy all have been shown to provide transposition function to an independent Mu element. Because Mu-A2 and Mu-R1 were isolated by analysis of the same a1-mum2 allele, and because molecular analysis showed these two elements have identical restriction maps (Chomet et al., Genetics 129:261-270, 1991; Qin et al., Genetics 129:845-854, 1991; this report), they most likely represent the same regulatory locus. Comparison of the nucleotide sequence of Mu-A2 with that of Mu9, a Mu element isolated from the bz2::mu9 allele (Hershberger et al., Proc. Natl. Acad. Sci. USA 88:10198-10202, 1991) reveals the near-identity of these two elements. Mu-A2 and Mu9 differ in sequence at only three nucleotide positions, suggesting that these two elements are the same and implying that Mu9 also is capable of providing transposition function to Mu1. Northern analyses of both Mu-R1 and Mu9 identified transcripts similar in size to those derived from Mu-A2. For reasons that are not clear, discrepancies exist in the reported sizes of the large transcript (3.4kb for Mu-A2; 2.9kb for Mu9, 2.5kb for Mu-R1). Differences in RNA size estimation may explain some or all of these discrepancies. Taken together, the data suggest that Mu-A2, Mu-R1, and Mu9 are nearly identical transposons capable of the autonomous regulation of Mu1. The relationship of these regulatory loci to Mu elements other than Mu1 remains to be discovered. However, because the family of Mu elements shares homology in regions of the terminal inverted repeats, it is predicted that the regulatory function of Mu-A2 will extend to the entire transposon family.
Figure
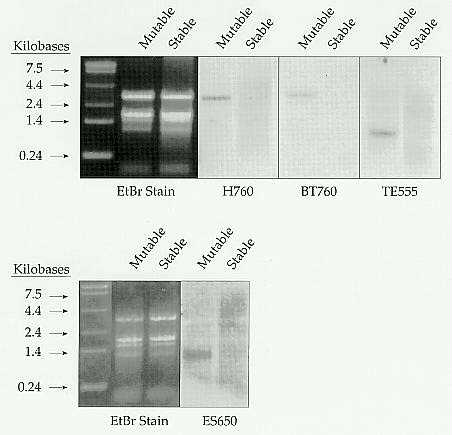
2. Transcription of Mu-A2. Total RNA isolated from seedlings
of mutable or stable a1-mum2 progeny in a cross showing 1:1 segregation
for the mutability trait was separated by electrophoresis in denaturing
agarose gels. Mu-A2 transcripts were detected using the indicated
restriction fragments (see Fig. 1) as random-primer labeled probes in Northern
hybridization analyses. In each case, transcripts were detected only in
RNA from mutable plants. Molecular weight standards are the 0.24-9.5Kb
ladder from Gibco-BRL.
Return to the MNL 66 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}