--John R. Laughnan, Susan Gabay-Laughnan and Janet M. Day
Evidence supporting the transposability of the naturally occurring restorer, Rf(CE1), in inbred line CE1, was reported in MNL 63:121 and has been extended in a companion article on these pages titled "Transposition characteristics of the naturally occurring cms-S restorer in inbred line CE1".
We present here preliminary evidence that plants that carry a transposed Rf most often retain Rf at or near the original (donor) site, a situation often referred to as replicative transposition. The system used to identify instances of Rf transposition involves the testcross: (S) rf rf wx wx X (S) Rf N Wx/rf T wx. The female parent on the left carries S-type cytoplasm, is phenotypically male-sterile (rf rf) and is homozygous for the mutant waxy-1 (wx) allele on chromosome 9. The male parent also carries S-type cytoplasm, is heterozygous for the restorer (Rf) to be tested for transposition capability, is heterozygous at the waxy locus (Wx wx), and for a translocation (T/N) chosen to effect a suitably close linkage between the particular Rf-gene, at what we will call the original, or control, site, and the wx locus. In these studies, map distances between Rf and wx in the translocation heterozygotes are around 12 map units or less. Since the restorers of cms-S, unlike those of cms-C and cms-T, act gametophytically, rf (nonrestoring) pollen grains of this male parent abort, so only Rf pollen grains function. Therefore, wx is transmitted through pollen only if there is a crossover between Rf and Wx to produce a functional Rf-wx microspore. On testcross ears that exhibit both Wx (starchy) and wx kernels, the Rf-wx recombination rate is given simply by the percentage of wx kernels, without the need to score progeny plants for the presence (male-fertile) or absence (male-sterile) of Rf. Instances of putative transposition are identified as male parents whose testcross ears show significantly enhanced frequencies of wx kernels. Many such cases of plants with enhanced Rf-wx recombination have been encountered in the natural restorers Rf(CE1) and Rf(Tr), and in the spontaneously occurring Rfs, I, III, IV and VI. Such higher Rf-wx recombination rates range from slightly above control rates, through intermediate levels, to rates exceeding 50%.
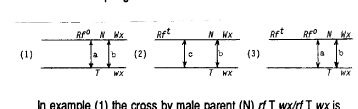
The evidence in support of replicative transposition of Rf genes is based on the frequency of male-fertile vs. male-sterile offspring from the cross (see Table 1 legend): (S) Rf N Wx/rf T wx X (N) rf T wx/rf T wx that is used to propagate the Rf heterozygotes whose testcrosses have been described above. When Wx kernels from this cross are planted, most of the progeny plants will be: (S) Rf N Wx/rf T wx heterozygotes with the same genotype as the female parent. If, in the female parent, a crossover occurs, either between Rf and T, or between T and Wx, the chosen Wx kernels, of either (S) rf N Wx/rf T wx or (S) rf T Wx/rf T wx genotype, will produce only male-sterile progeny. The three simple diagrams below illustrate heterozygous female parents of three possible types: (1) a single Rfo at the original, or control site, (2) a single transposed Rft at a recipient site some distance from the Rfo site, and (3) two Rf genes, one (Rfo) at the original site, and the other (Rft) at the transposition site. The arrows indicate crossovers that can yield Wx-non-Rf male-sterile recombinant offspring.
In example (1) the cross by male parent (N) rf T wx/rf T wx is expected to produce from 5 to 10% Wx-non-Rf male-sterile progeny corresponding to the short map distance between Rfo and Wx (crossovers a and b).
In example (2) either crossover b or c will produce Wx-non-Rf male-sterile progeny; since in this case the Rft-Wx distance is identifiable, from independent testcrosses, as significantly above the control level, and may range up to and above 50%, the frequencies of male-sterile (Wx-non-Rf) offspring are expected to be proportional to the Rft-Wx distance and should approach levels of 50% of offspring in some instances.
In example (3), where Rfo and Rft are both present, only crossovers a and b will yield Wx-non-Rf male-sterile offspring, since a crossover between Rft and Rfo results in a Wx-Rfo male-fertile offspring. Thus, given a female parent that has been shown by independent testcrossing to carry a transposed Rft, the progeny of its cross with the (N) rf T wx/rf T wx male parent should exhibit a male-sterile frequency corresponding to the Rft-Wx distance if Rfo is no longer present at the original site (nonreplicative). If, however, Rfo is present at its original site (replicative), only control frequencies of male-sterile offspring are expected, no matter how distant Rft may be from Rfo, and this holds even if Rft is located in a nonhomologous chromosome.
The data in Table 1 provide a test of the above expectations. In column 1 are given the genotypes of four different Rf heterozygotes in the study. For example, in the case of the Rf(CE1) N Wx/rf T wx heterozygote (row 1), there were 30 plants involved as female parents in the indicated cross. On the basis of the performance of each of these plants as male parents in testcrosses into (S) rf rf wx wx testers, 14 (column 3) gave relatively low Rf-wx recombination rates, from 3.8% to 14.8% (column 2). Columns 4-7 (row 1) deal with the distribution of male-fertile and male-sterile progeny from crosses of (S) Rf(CE1) N Wx/rf T wx plants as female parents with (N) rf T wx/rf T wx testers. Sixteen (column 9) of the 30 Rf(CE1) heterozygotes, when used as male parents in testcrosses, gave Rf-wx recombination from 16.0% to 45.4% (column 8, row 1); the distribution of male-fertile and male-sterile offspring from these 16 Rf(CE1) heterozygotes is given in columns 10-13, row 1. Because of their higher levels of Rf-wx recombination these 16 heterozygotes are regarded as Rf transpositions. Corresponding data for RfI and RfIII heterozygotes are given in Table 1. In the case of RfVI, only higher-level recombination heterozygotes were available to score.
Table 1. Frequencies of male-sterile offspring from (S) Rf N
Wx/rf T wx* X (N) rf T wx/rf T wx crosses,
arranged according to Rf-wx recombination rates in progeny of testcrosses
of the female parent.
| Genotype of restorer parent | From crosses whose female parents gave lower-level Rf-wx recombination rates in testcross | From crosses whose female parents gave higher level Rf-wx recombination rates in testcross | ||||||||||
| % Rf-wx recombination | No. of crosses | male-fertile progeny | male-sterile progeny | Total progeny | % male-sterile | % Rf-wx recombination | No. of crosses | male-fertile progeny | male-sterile progeny | Total progeny | % male-sterile | |
| Rf(CE1) N Wx/rf T2-9d wx | 3.8-14.8 | 14 | 99 | 8 | 107 | 7.5 | 16.0-45.4 | 16 | 92 | 10 | 102 | 9.8 |
| RfI N Wx/rf T8-9 (043-6) wx | 3.0-11.0 | 13 | 117 | 6 | 123 | 4.9 | 24.5-33.0 | 7 | 107 | 3 | 110 | 2.7 |
| RfIII N Wx/rf T2-9d wx | 0.0-6.2 | 13 | 114 | 5 | 119 | 4.2 | 32.8-56.5 | 6 | 74 | 1 | 75 | 1.3 |
| RfVI N Wx/rf T2-9d wx | -- | -- | -- | -- | -- | -- | 39.4-69.5 | 3 | 43 | 3 | 46 | 6.5 |
| TOTALS | 40 | 330 | 19 | 349 | 5.4 | 32 | 316 | 17 | 333 | 5.1 | ||
The data provide no evidence for higher levels of male-sterile plants among the progeny of Rf heterozygotes with high Rf-wx recombination rates. The frequencies of male-sterile plants among offspring of Rf heterozygotes in low and high recombination categories are remarkably similar in Rf(CE1), RfI and RfIII restorer strains and all are at control-rate levels. In the case of RfVI, for which data are available only for the high Rf-wx recombination category, the male-sterile rate (6.5%) is also at the control level. Overall, a total of 349 plants from Rf heterozygotes with low Rf-wx recombination rates, and 333 plants from Rf heterozygotes with high Rf-wx recombination rates were scored. The frequencies of male-sterile offspring in these two groups were 5.4% and 5.1%, respectively. The contingency chi-square for these data corresponds to a P value of 0.85.
It is evident that an overwhelming proportion of plants with enhanced
Rf-wx recombination rates (transpositions) retain an Rf at
or near the original Rfo site. This is not to say that, with the
use of other crossing protocols, Rfo can not be removed through
recombination; we are currently studying this possibility.
Return to the MNL 66 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}