--Ming T. Chang
A project was conducted in 1988 aimed at rapidly generating pure inbreds through double haploids. These double haploids are genetically induced by Stock 6, a line which has been reported to generate a high frequency of haploids. Two inbreds that were derived from Oh43 and Mo17 were crossed and the F1 seeds were used as female parent. The genetic constitution of the hybrid is homozygous A b pl c1 r-g Y. So the hybrid kernels have a deep yellow color and the plants have green leaves, sheath, tassel and anther. The male is a BC2S2 that was derived from crosses between Stock 6 (A b pl C R-g y) and A B Pl C R-nj y genetic stock. An isozyme test showed that the BC2S2 material was still segregating at Adh1 and Pgm2 loci (see above article), but 90% of the individuals were fixed at those loci.
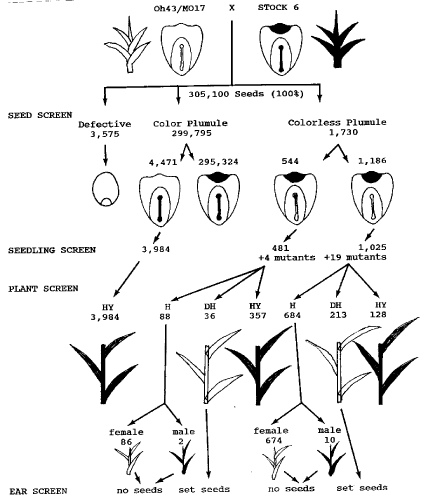
A total of 600 ears (305,778 seeds) were obtained from crosses between the F1 hybrid and BC2S2 Stock 6. In theory, the seeds on the ear should all be purple crown-purple plumule (due to R-nj) and light yellow (due to 2 doses of Y). But some kernels were colorless (due to contamination or development of the unfertilized endosperm). The contaminant kernels should be colorless (due to r-g) and deep yellow (due to 3 doses of Y), and could be easily identified. Using this endosperm color screening technique, the contaminated kernels were first removed from the ear. A total of 678 contaminants (0.22%) were identified, of which 75 kernels had purple plumule color, the remaining 603 kernels had colorless plumules. These 75 kernels were the result of heterofertilization. One sperm from Stock 6 pollen fertilized the egg and one sperm from F1 pollen fertilized the polar nuclei.
Among the remaining 305,100 kernel samples, 4,471 kernels (1.5%) had colorless endosperm and colored plumule, and 3,575 kernels (1.2%) were defective (Figure 1). These 4,471 colorless kernel samples had light yellow endosperm (due to 2 doses of Y) and plants were purple in color. Kernel size was slightly reduced but the difference was not significant. These kernels should be derived from the development of fertilized egg with unfertilized endosperm, although the chromosome ploidy of endosperm tissue was not determined. The defective kernels were not able to germinate. The origin of these defective kernels was not known.
The remaining purple crown (due to R-nj) seeds were then screened for embryo plumule color. Among those seeds, we discarded 295,324 (96.8%) which had colored plumule. 1,730 (0.57%) seeds that had colorless plumule (Figure 1) were planted and at 2 to 3 leaf stage, a 0.05% colchicine solution was applied. A total of 23 mutants were obtained from these seedlings. Among those mutants were 3 albino, 7 white stripe, 1 dwarf, 4 yellow green, 5 pale green, 2 necrotic crossband, and 1 tiny seedlings. The surviving 1,506 plants were grown to maturity. Among those, 485 plants proved to be purple hybrids due to crosses between the F1 hybrid and Stock 6. However, 249 green plants did set yellow seeds (presumed to be double haploids), and the remaining 772 plants were 100% sterile (presumed to be haploids). The 772 haploid plants were further classified into two types, 760 small sterile green plants and 12 small sterile purple plants. Root tip examination from 10 plants showed that they were all haploids with 10 chromosomes. RFLP examination from another 10 plants showed a single band at all ten loci. It is therefore concluded that these small seedlings are true haploids. The parthogenetic and androgenetic ratio from Stock 6 is 760 vs. 12, or 98.45% vs. 1.55%.
Figure 1. A schematic procedure for double haploid production using Stock 6. H = haploid, DH = double haploid, HY = hybrid.
The homozygosity of these 249 putative double haploid lines was examined by isozyme analysis. Ten seedlings from each line were examined for 10 different isozyme loci. Among those, 98 lines were proved to be homozygous pure inbred lines. The remaining 151 lines were segregating at 1, 2, or 3 isozyme marker loci as shown in Table 1. Results clearly showed that these plants were not derived from Stock 6. It was clear from isozyme analysis that these plants came from self-contamination. Since they had light yellow (due to y) and purple crown (due to R-nj) endosperm, it was evident that these seeds were the result of heterofertilization. The embryo was selfed and the endosperm was fertilized by Stock 6. All 249 lines were planted ear to row in 1989. Morphological uniformity was evaluated. Based on isozyme and morphological data, only 98 lines were pure inbreds and the rest were hybrids.
Table 1. Isozyme analysis of the putative double haploids.
| Genetic Constitution | |||||
| Material | Frequency | Mdh1 | Mdh2 | Pgd1 | Pgd2 |
| Stock 6 | 22 | 22 | 22 | 22 | |
| Oh43 derivative | 11 | 11 | 11 | 22 | |
| Mo17 derivative | 22 | 22 | 11 | 11 | |
| Hybrid x Stock 6 | 23 | 11 | 12 | 11 | 12 |
| 26 | 22 | 12 | 11 | 12 | |
| 25 | 12 | 11 | 11 | 22 | |
| 20 | 12 | 22 | 11 | 11 | |
| 57 | 12 | 12 | 11 | 12 | |
The number of haploids now equals 772 haploids + 98 double haploids
= 870, or 0.29% (870/305,100). That means 0.29% of the pollen grains preferentially
fertilized the polar nuclei. If all of the 4,471 (1.47%) colorless endosperm
colored plumule kernels are diploid embryos with unfertilized diploid endosperms,
then the pollen grains must have preferentially fertilized the egg first.
Thus, if lack of fertilization is the cause of haploidy, then when a single
sperm goes inside the embryo sac the chance of fertilizing the egg first
is 83.71%, and the chance of fertilizzing the polar nuclei first is 16.29%.
It is therefore concluded that a single sperm alone will preferentially
fertilize the egg.
Return to the MNL 66 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}