Correlation Genetics
URBANA, ILLINOIS
University of Illinois
A repeated sequence derived from the Arabidopsis telomere is dispersed in the maize genome as detected by in situ hybridization
--Carol M. Uyeda, Manju Gupta and Dale M. Steffensen
We were interested in obtaining DNA markers adjacent to each of the twenty maize telomeres in order to define the physical ends of the chromosome arms. To accomplish this, we cloned high-molecular weight maize genomic DNA into a selective yeast artificial chromosome (YAC) vector that contains only one telomere. The other telomere would be supplied by maize chromosomes by complementation in yeast. As a result, only those yeast clones that contained both telomeres would propagate in yeast. A chromosome end-specific YAC library of 8,000 clones was generated and screened with an oligonucleotide probe (CCCTAAA)6 derived from the Arabidopsis telomeric sequence (ATS). Positive clones were tested for Bal31 sensitivity, which showed whether the ATS was present at the YAC terminus. The clones showing localization of ATS hybridizing sequences at the ends of the YACs were selected for further work.
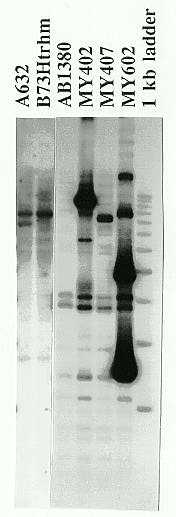
To see whether the sequences in the YAC clones that hybridized to the ATS were actually maize telomeres, they would have to be subcloned and used as hybridization probes to maize chromosomes in situ. To bypass the subcloning procedure, we adapted the following strategy: total DNA of yeast strain MY602, which contained a putative telomeric YAC clone, was amplified with a single primer, 5'-d(TTTAGGG)-3', resulting in a few amplified bands. No bands were amplified with this primer from genomic DNA of the yeast strain AB1380, which did not contain any YAC. The amplified DNA from MY602 was fractionated in an agarose gel, the DNA was denatured and transferred to a nylon membrane. The blot was then probed with the ATS. The most strongly hybridizing fragments of 0.7kb and 1.5kb were isolated and used to probe a Southern blot of digested maize (A632 and B73 Ht rhm) and yeast (control AB1380, MY602, MY402 and MY407) genomic DNA (Figure 1). The 1.5kb fragment hybridized strongly to one band in B73 Ht rhm and to two bands in A632. The probe also hybridized strongly to a few fragments of MY602 DNA which were not present in the AB1380 DNA. This demonstrated that the probe sequence was not specific to yeast genomic DNA, and that it originated from maize DNA sequences cloned in the YAC in MY602. The other amplified fragment of 0.7kb produced the same patterns. The 1.5kb fragment was subcloned in a plasmid vector and the purified insert was labeled with 35S-substituted dATP to a specific activity of ~1 X 108 dpm/ug using the Stratagene Prime-It DNA labeling kit. This labeled fragment was used as a probe for in situ hybridization.
Figure 1. Hybridization of the labeled 1.5kb fragment to EcoRI-digested total DNA from maize and yeast. The probe came from the amplification of total DNA of yeast strain MY602 with the Arabidopsis telomeric repeat primer. For reference, the smaller of the two most prominent bands in lane #1 (A632) is approximately 8kb in size.
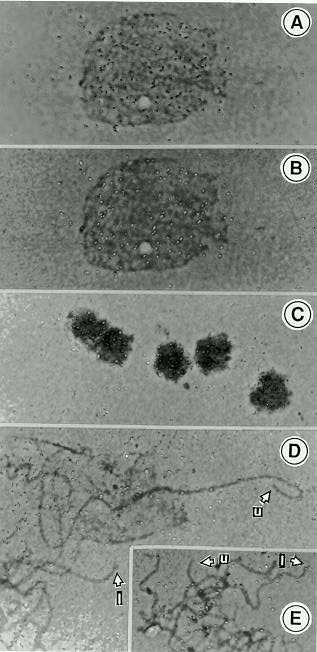
As demonstrated by in situ hybridization to maize chromosomes, the MY602 derived sequence appeared to be distributed over the maize genome with no particular localization at the telomeres. Several of the telomeres (shown in Figure 2D and 2E) were not detectably labeled, while others were. The labeled regions appeared to be randomly distributed over chromosomes and nuclei with no definitive "hot spots", as would be expected for a medium repetitive DNA sequence. The inbred line B73 was quite satisfactory for this study, especially when sporocytes were used to make slides immediately after fixation in 95% ethanol:glacial acetic acid, 3:1 (v/v). Unlike older material, freshly fixed meiotic nuclei often popped free of their cell walls. Cell walls will bind probes non-specifically, thus obscuring the analysis. In nuclei (Figure 2A, 2B, 2C) the silver grains from the probe were distributed evenly and not at the edges where the telomeres would be attached to the nuclear membrane. The result shows that 35S provides good enough resolution to decide whether or not the hybridization is on the long arm, the short arm, or at the telomeres of the chromosomes. Our calculations show that a specific activity of at least 1 X 108 dpm per ug is sufficient to detect single copy sequences of 1-2kb in length.
These results show that the 1.5kb DNA fragment from the MY602 YAC is a middle repetitive sequence, which is not only present in the subterminal regions of the chromosomes but is also dispersed throughout the maize genome.
Figure 2. In situ hybridization with a 35S-labeled putative maize telomere sequence: A and B are the same early meiotic nucleus photographed at two levels of focus. In A the focus is on the silver grains (black). In B the focus is on the nucleus, where the silver grains show in a refractive white color (the same in C, D, and E). Panel C shows uniformly labeled tapetal nuclei. D and E are labeled pachytene chromosomes. Unlabeled telomeres are identified as u, and labeled telomeres as l.
B73 tassels at meiotic stages were fixed in 3:1 (v/v) ethanol:acetic
acid and slides of sporocytes were prepared as squashes in 45% acetic acid,
frozen, dehydrated in ethanol and air dried. The chromosomal DNA on the
slides was melted in 70% formamide at 70 C. The double-stranded labeled
DNA probe in 50% formamide, 2X SSC and 1mg/ml E. coli tRNA as carrier
was heated to 70 C and quick-cooled on ice. Hybridization was at 35 C.
Slides were dipped in Kodak NTB2 liquid emulsion and exposed for either
a week or a month. They were then stained with Giemsa and photographed
under phase contrast.
Return to the MNL 66 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}