University of Minnesota
MADISON, WISCONSIN
University of Wisconsin
Teosinte glume architecture (tga1), a locus from teosinte
-- John Doebley, Jane Dorweiler and Jerry Kermicle
The glumes of the maize ear are normally (1) relatively soft, (2) oriented perpendicular to the axis of the ear, i.e. reflexed, (3) relatively short such that the palea and lemma are readily visible on a shelled cob, and (4) shovel-shaped. In contrast, the glumes of the teosinte ear are (1) highly indurated, (2) oriented parallel to the axis of the ear, i.e. erect, (3) relatively long such that they cover the palea and lemma, and (4) boat-shaped. We refer to the former complex as maize glume architecture (MGA), and, correspondingly, the latter complex as teosinte glume architecture (TGA).
In the process of transferring cross-incompatibility loci on 4S from teosinte into maize inbred W22 (Kermicle, J and Allen, Maydica 35:399-408, 1990), a factor producing TGA was also transferred into this inbred. In W22 background, the TGA trait is stably expressed. During six generations of backcrossing to W22, the TGA trait behaved like a single locus. In W22 background, TGA is essentially recessive to MGA. Heterozygotes are somewhat intermediate, but much more closely approach the standard W22 phenotype.
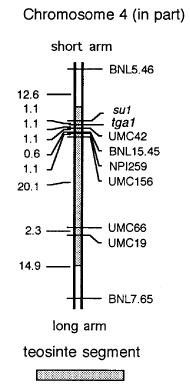
We attempted (1) to test if the factor transferred from teosinte to maize would segregate as a single locus and (2) to map its chromosomal location. First, we compared W22 to W22-TGA (W22 converted to TGA) for their genotypes at a series of molecular marker loci on chromosome 4. Molecular marker loci for which these two lines carried different alleles should lie within the chromosome segment transferred from teosinte. The two lines should possess the same allele at all marker loci outside the transferred segment. It was also known that the W22-TGA lost one of the teosinte incompatibility loci that lies 4 map units distal to su1 during the backcrossing program. Using these two sources of information, we determined the extent of the segment of teosinte chromosome 4 transferred into W22 (Fig. 1). The segment is minimally 32 map units in length.
We also crossed W22-TGA to a su1 tester and then backcrossed an F1 plant to W22-TGA. One hundred-seventy BC1 plants were grown and classified for glume architecture. In addition, nine molecular marker loci and su1 were scored in the BC1 plants. Results of this experiment did not tabulate as neatly as originally envisioned. Rather than segregating into two discrete classes (TGA and MGA), a continuous bimodal distribution of glume architecture phenotypes was observed. Thus, we classified plants as either MGA, TGA or intermediate. Sixteen percent (27 plants) of the plants had an intermediate phenotype. We also classified individuals which showed either clear TGA or MGA as having moderate or strong expression of the trait (Table 1).
Figure 1. Linkage map of a portion of chromosome 4 showing the position of tga1 and the marker loci used to localize it. The position of tga1 should be considered approximate until mapped in a background in which it is stably expressed.
Table 1. Classification of BC1 plants for either teosinte (TGA) or maize
(MGA) glume architecture and the degree of expression of the trait.
| Degree of Expression | |||
| Trait | Moderate | Strong | Total |
| TGA | 39 | 29 | 68 |
| MGA | 49 | 26 | 75 |
We considered two possible hypotheses to explain the failure of the
glume architecture trait to segregate cleanly. (A) The segment transferred
from teosinte contained multiple loci affecting glume architecture and
the intermediate class represents individuals with some but not all of
these linked loci because of cross-overs within the segment. (B) Glume
architecture is largely controlled by a single major locus within the transferred
segment, but differences in background (other unlinked loci) between W22
and the su1 tester affect the expression of this putative major
locus, giving rise to the intermediate phenotypes. To distinguish between
these two possibilities, we examined the amount of recombination between
the marker loci for plants in the intermediate phenotype and strong expression
classes. If hypothesis (A) is correct then the intermediate individuals
should all contain cross-overs within the chromosome segment transferred
from teosinte. If hypothesis (B) is correct then the number of cross-overs
in this segment should be the same for the intermediate and strong expression
classes.
Examination of the data clearly supported hypothesis (B). First, ten of the 27 (44%) plants with intermediate phenotypes contained no cross-overs in the entire transferred segment from BNL5.46 to BNL7.65. Thus, the intermediacy of these plants is not the result of recombination between loci in the transferred segment (hypothesis A). Second, the distribution of cross-overs within the segment in these two groups is equivalent, and the intermediate class does not show significantly greater recombination between any pair of adjacent marker loci (Table 2). Thus, there is no evidence to suggest that the intermediate phenotypes result from break-up of linked loci within the segment transferred from teosinte. Rather, it appears that unlinked factors affect the degree of expression of the glume architecture traits in our population.
Table 2. Percentage of chromosomes with cross-overs between adjacent
marker loci in the full population (FP), and in the subpopulations with
strongly expressed glume architecture traits (SE) and intermediate phenotypes
(IP).
| Marker Loci | FP | SE | IP |
| BNL5.46 - su1 | 12.6 | 12.7 | 18.5 |
| su1 - UMC42 | 2.3 | 1.8 | 0.0 |
| UMC42 - BNL15.45 | 1.1 | 0.0 | 3.7 |
| BNL15.45 - NPI259 | 0.6 | 0.0 | 0.0 |
| NPI259 - UMC156 | 1.1 | 1.8 | 0.0 |
| UMC156 - UMC66 | 20.1 | 18.2 | 22.2 |
| UMC66 - UMC19 | 2.3 | 5.5 | 0.0 |
| UMC19 - BNL7.65 | 14.9 | 18.2 | 14.8 |
Based on these results, we designate a locus tga1 (teosinte glume
architecture). In W22 background, tga1 from teosinte produces the
phenotype described above as TGA. Our data indicate that tga1 resides
in the interval between su1 and UMC42, approximately 1 map
unit from su1 (Fig. 1). To further investigate these results, we
will map tga1 in a population derived from a cross of W22-TGA with
W22. In this population, there will be no background effects and thus all
progeny should be readily classified. As discussed elsewhere (Doebley et
al., PNAS 87:9888-9892, 1990), this locus on chromosome 4 is one of the
key loci differentiating maize and teosinte.
Return to the MNL 66 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}