--L. Bernard, P. Ciceri, B. Lazzari and A. Viotti
The maize o2 locus codes for a DNA-binding protein which activates the transcription of a group of zein genes (Schmidt et al., Proc. Natl. Acad. Sci. 87:46-50, 1990). This protein belongs to the b-Zip family of transcriptional activators, like the jun and fos mammalian oncogenes, the yeast GCN4 and the plant TGAI and HBPI (Tabata et al., EMBO J. 10:1459-1467, 1991; Busch and Sassone-Corsi, Trends Genet. 6:36-40, 1990; Katagiri et al., Nature 340:727-730, 1989). The activation is determined by the binding to short target sequences (Lohmer et al., EMBO J. 10:617-624, 1991; Hoscheck and Schmidt, unpublished) upstream to the CAAT and the TATA boxes, which are present in almost all the high molecular weight zein genes (H) and usually absent in the low molecular weight genes (L).
The hybridization of Southern and Northern filters with O2 probes (kindly provided by R. Schmidt, R) showed a high level of heterogeneity in a number of o2 maize lines for what concerns the DNA restriction pattern and the type and level of transcription products.
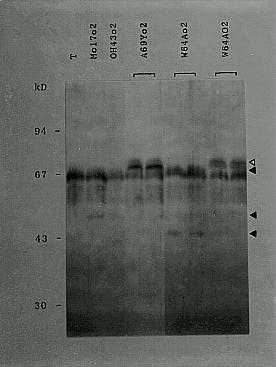
Figure 1. Western blot of several maize lines using a polyclonal O2 antibody. T = total protein extract from tobacco leaves identifying the antibody cross-reactions bands. D = O2 double band. s = truncated O2 proteins.
The Northern results are supported by the Western analysis of total endosperm proteins (carried out with O2 antibodies), where the different o2 lines show the absence or the presence of O2 modified polypeptides (Fig. 1). It has in fact been possible to identify a double band in all the examined wildtype lines (W22, A69Y, NYR and W64A) at the apparent O2 wildtype molecular weight region (68 and 69kDa), that may represent products of post-translational modification (e.g. phosphorylation or O-glycosylation) of the same protein. These two bands are always absent in the o2 mutations (W22 o2, A69Y o2, NYR o2 and W64A o2) where we can detect O2 transcripts and notice the presence of a number of specific bands of different molecular weight which probably are truncated or unmodified O2 polypeptides. In the two lines OH43 o2 and Rossman o2 that are null-transcript, there is no O2 specific extra-band and they can be considered as negative controls.
The timing analysis of O2 expression during seed development (at 14, 20 and 25 DAP) shows, on the basis of the same amount of total RNA or proteins, a maximum of transcription and translation at 14 DAP. Furthermore, it has been observed by Northern analysis and SDS-PAGE that the various o2 mutations differentially affect the transcription and the product level of both the H and L zein genes.
These results suggest that the O2 mutated proteins may specifically
affect the expression of the various subsets of the zein multigene family,
according to the kind of modification they encounter.
Return to the MNL 66 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}