Agrigenetics Company
URBANA, ILLINOIS
University of Illinois
WASHINGTON, DC
Smithsonian Institution
Another cytological marker on the Agrigenetics maize RFLP linkage map
David Zaitlin, Dale M. Steffensen and Elizabeth A. Zimmer
In maize and its close relatives, the more than ten thousand copies of the cytoplasmic ribosomal RNA genes (18S-5.8S-26S RNAs) are organized into a large tandemly repeated cluster on chromosome 6 (Phillips, RL et al., Stadler Symp. 15:105-118, 1983; Zimmer, EA et al., Genetics 120:1125-1136, 1988). This structure, known as the nucleolar organizer region (NOR), is a familiar cytological feature of the short arm of chromosome 6 and is the most distal marker on 6S on several maize genetic maps based on restriction fragment length polymorphisms (RFLPs) (see Burr, B and Burr, FA, Trends Genet. 7:55-60, 1991). Cytological and genetic studies (Mascia, P et al., MNL 54:112, 1980; Mascia et al., Gene 15:7-20, 1981; Steffensen, DM and Patterson, EB, Genetics 91:s123, 1979) have established that the genes encoding the 5S rRNA of maize are also clustered in a large repeating array of several thousand copies/haploid genome on the long arm of chromosome 2. The cytological site of the 5S genes was bracketed between two translocation breakpoints (0.75 to 0.82) using the translocations T2-4f (2L.75:4L.12) and T2-6(5419) (2L.82:6S.79). The 5S rRNA genes have been placed on the RFLP map of tomato at a single site on chromosome 1 (Lapitan et al., Genome 34:509-514, 1991). Here we show that sequence polymorphism exists between various maize inbred lines within or around the 5S gene cluster, and that standard RFLP mapping methodology identifies a single genetic locus on 2L between the RFLP loci BNL6.20 and C2b.
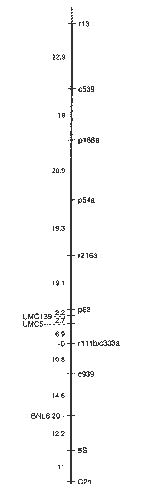
The 322bp BamHI 5S repeat fragment from pZm5S5 (Chen and Zimmer, EA, Fed. Proc. 43:1557, 1984) was subcloned into pJKKmf(-) (Kirschman and Cramer, Gene 68:163-165, 1988) to enable the synthesis of a single-stranded 32P-labelled RNA probe with either SP6 or T7 RNA polymerase. Initial screening of EcoRI digests of total DNA extracted from a group of maize inbreds revealed that the probe hybridized primarily to a diffuse region on the blots at the upper limit of resolution (>30kb in length). However, there were also eight discrete bands present of somewhat lower signal intensity, ranging between 6kb and approximately 26kb in length. Six of these bands were common to both B73 Ht rhm and B68 Ht. Bands of ~23kb and 10.3kb were unique to B73 Ht rhm and B68 Ht, respectively. The 5S RNA probe was hybridized to mapping blots containing EcoRI-digested samples of genomic DNA extracted from 93 F2 individuals from the cross B68 Ht X B73Ht rhm. The two diagnostic bands of ~23kb and 10.3kb were scored individually for presence vs. absence. Linkage analysis showed that the two bands mapped to the same locus (therefore they are allelic), and the autoradiograms were rescored to reflect this. The 3-class 5S rDNA data were incorporated into a large data set containing the allelic scores for 94 RFLP probes that were polymorphic in this population with either EcoRI or EcoRV. The 5S rDNA cluster showed linkage to 5 probe loci, all from chromosome 2, using the chi-square goodness-of-fit test at a significance level of p=0.05. The Agrigenetics program Surveyor was used to order the markers and minimize the map distances. The resulting linkage map is shown in Figure 1.
Figure 1. RFLP linkage map of chromosome 2. All maize RFLP probes were mapped in a population consisting of 93 F2 progeny from the cross B68 Ht X B73Ht rhm. Agrigenetics probe loci are shown on the right side of the figure and loci detected by UMC and BNL probes are on the left. The figure is oriented with the short arm of the chromosome at the top (the position of the centromere was not determined). Linkage distances are given in percent recombination, and the location of the 5S rDNA gene cluster is shown in boldface.
Without further investigation we can only speculate as to how the accessory bands that we scored in the mapping of the 5S rDNA gene cluster are generated by EcoRI (a few of these bands are just visible in Figure 4 of Mascia, P et al., Gene 15:7-20, 1981). Because they hybridize strongly to the 5S RNA probe relative to known single-copy sequences, these bands probably represent DNA fragments that individually contain many (between 20 and 75) copies of the 5S repeat unit that arise within the cluster itself or at the junction(s) of the cluster and adjacent sequences. We find that EcoRI is much less sensitive to the presence of 5-methylcytosine in plant genomic DNA than are some other restriction enzymes (EcoRI will digest sunflower and maize DNA to completion, whereas PstI and SalI will not). Effects attributable to methylation have not been observed with EcoRI in the construction of RFLP maps using hundreds of maize DNA clones in several different populations. This would, therefore, tend to rule out such heritable chemical modification as the source of genomic polymorphism between B68 Ht and B73 Ht rhm. Examination of the nucleotide sequence of the cloned maize 5S rDNA repeat (Figure 2) reveals that there are three subsequences, all within the coding region of the 5S RNA (GAACTC at positions 41-46, GAAGTC at positions 99-104, and GCATTC at positions 113-118) that could each become recognition sites for EcoRI (5'-GAATTC-3') with a single base change. Gain of an EcoRI site (due to an actual base change, not methylation) has been observed in 10%-50% of the repeat units of the rDNA array in some inbred lines of maize (Zimmer, EA et al., 1988); Jupe and Zimmer, Plant Mol. Biol. 14:333-347, 1990). Conceivably, such mutational events could also have occurred within the 5S gene cluster. Mascia, P et al. found that a small percentage of the BamHI sites located within the coding region (nucleotides 30-35) were no longer cleaved by this enzyme due to sequence divergence, although none of these variants were sequenced. As few as nine EcoRI sites could account for the eight bands seen on the autoradiograms (given that there is an EcoRI site between each end of the cluster and the adjacent sequences), although this is certainly a minimal estimate.
Figure
2. Nucleotide sequence of the 322bp maize 5S rRNA repeat from pZm5S5.
The 120bp coding sequence of the 5S RNA is shown in upper case letters
and the sequence of the spacer region is shown in lower case. Conserved
recognition sites for BamHI and TaqI are underlined, and
the three potential EcoRI recognition sites are emboldened. Comparisons
of the maize 5S rDNA sequence with those from five other plant species
reveals that the subsequences GAACTC (41-46) and GAAGTC (99-104) are conserved
in the genes from wheat, flax, yellow lupine, rye and rice, while the subsequence
GCATTC at positions 113-118 is present only in maize.
Return to the MNL 66 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}