Agrigenetics Company
SAN JOSE, CALIFORNIA
Correlation Genetics Co.
Linkage of a second gene for NCLB resistance to molecular markers in maize
--David Zaitlin, Sandra J. DeMars and Manju Gupta
There are presently four dominant or partially dominant genes known that individually condition resistance in maize to Exserohilum turcicum, the northern corn leaf blight (NCLB) pathogen. These genes, known as Ht1, Ht2, Ht3 and HtN, can be readily differentiated from one another by the reaction they confer in maize to the known physiologic races of E. turcicum. Plants homozygous or heterozygous for either Ht1, Ht2 or Ht3 express a qualitative form of resistance characterized by lesions that are extensively chlorotic, rather than wilted and necrotic, and within which fungal sporulation is suppressed (Smith and White, Disease of Corn, in Corn and Corn Improvement, pp. 687-766, 1988). HtN, in contrast, acts quantitatively by delaying the onset of lesion development until after flowering (Gevers, HO, Plant Dis. Rep. 59:296-299, 1975). Ht1 was identified nearly 30 years ago in the inbred GE440 and a Peruvian popcorn introduction (Hooker, AL, Phytopath. 53:660-662, 1963; Ullstrup, AJ, Plant Dis. Rep. 47:107-108, 1963), and has long been known to reside on the long arm of chromosome 2 at position 121 between w3 and Ch1. Studies employing molecular markers have demonstrated correlations between the presence of this gene and RFLPs at several loci on 2L between resistant inbreds carrying Ht1 (introduced through backcrossing) and the corresponding susceptible parents (Hoisington, DA and Coe, EH, Jr., Curr. Commun. Mol. Biol., Cold Spring Harbor: 19-24, 1989; Chyi et al., MNL 63:112, 1989). The results of a genetic study linking Ht1 to RFLP loci on 2L can be found in Bentolila et al. (Theor. Appl. Genet. 82:393-398, 1991).
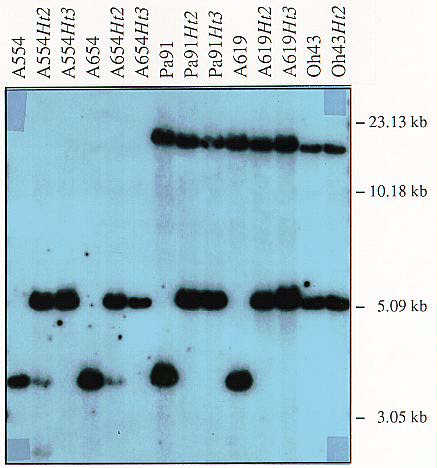
The chromosomal locations of Ht2, Ht3 and HtN have not been reported, although Ht2 (from NN14) has been shown to segregate independently of both Ht1 (Hooker, AL, Crop Sci. 17:132-135, 1977) and Ht3 (Simone, GW, Ph.D. dissertation, Univ. Ill.-Urbana, 1978; Hooker, MNL 55:87-88, 1980). While conducting a comprehensive survey of maize inbred lines, we observed RFLPs between restriction enzyme digests of genomic DNA from several near-isogenic inbred line (NIL) pairs that differed primarily at the Ht2 locus (Ht2/Ht2 vs. ht2/ht2) at two molecular marker loci on chromosome 8. The genomic clone UMC48 detected polymorphisms with EcoRI, HindIII and EcoRV between A654 and A654 Ht2, Pa91 and Pa91 Ht2, and A619 and A619 Ht2. The fragments to which the probe hybridized were of a unique size in each of the Ht2 lines; the actual size was dependent upon the particular restriction enzyme (see Figure 1). The same polymorphism was observed between A554 and A554 Ht2 with HindIII and EcoRV, between Oh43 and Oh43 Ht2 with EcoRI and EcoRV, and between Va26 and Va26 Ht2 with EcoRV only. UMC30, which is closely linked to UMC48, showed a similar correlation with EcoRI between A554 Ht2, A654 Ht2, Pa91 Ht2, A619 Ht2 and Va26 Ht2 and their recurrent counterparts, and between Pa91 and Pa91 Ht2 and A619 and A619 Ht2 with HindIII (EcoRV was not used).
Figure 1. Hybridization of 32P-labelled UMC48 to HindIII-digested DNA from fourteen different maize inbred lines. Molecular size standard values are shown on the right side of the figure. No polymorphism was observed between Oh43 and Oh43 Ht2 at this locus with this restriction enzyme. The high Mw band seen in Pa91, A619, Oh43 and the NCLB-resistant versions of these lines segregates independently of the other bands in our Ht2 mapping population, but remains unmapped (D.Z., unpublished).
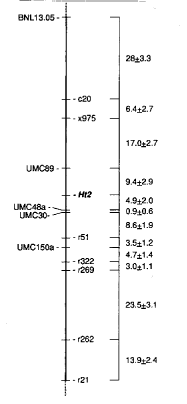
Approximately 375 F2 plants from the cross A619 Ht2 X W64A were inoculated with an aqueous conidial suspension of E. turcicum Race 1 in the field at Madison 6 weeks after planting. Chlorotic flecking, characteristic of early E. turcicum infection, was observed on all plants 4-5 days later. Weather conditions in the summer of 1991 were nearly ideal for NCLB disease symptom development, and large lesions were evident within three weeks of inoculation. The first 124 F2 plants showing clear, unambiguous symptoms of NCLB were rated and tagged (R:S=2.7:1). Genomic DNA was extracted from ear husk tissue taken from all 124 plants, digested with EcoRI, electrophoresed on 0.8% agarose gels and transferred to Genatran 45 nylon membrane. These mapping blots were then sequentially hybridized to twelve RFLP probes that were previously known to map to loci on chromosome 8. The allelic segregation scores for these markers (3-class) were analyzed together with the NCLB field disease ratings (2-class). Eight of the RFLP markers showed linkage to Ht2 (significance level p=0.05). Recombination frequencies between linked loci were determined by the method of maximum likelihood (Allard, RW, Hilgardia 24:235-278, 1956), and marker order and interlocus map distances (percent recombination) were determined using the method of minimization by simulated annealing (Metropolis et al., J. Chem. Phys. 21:1087, 1953). The linkage map is shown in Figure 2.
Figure 2. Genetic linkage map of chromosome 8 in maize. Linkage values are shown in percent recombination. Nomenclature for Agrigenetics probes (loci shown on the right) are: c=coleoptile cDNA clone, r=root cDNA clone, x=genomic XhoI clone. Probes with UMC and BNL prefixes are PstI-generated maize genomic clones originating from the University of Missouri at Columbia and Brookhaven National Laboratory, respectively.
This is the first report of the genetic mapping of Ht2 in maize.
While we consider this to be a preliminary study due to the use of F2 plants
in a field environment, the linkage arrangement of the RFLP markers provides
a strong internal control for population structure. We find that the order
and relative linkage distances agree quite closely with those determined
separately in three other independent mapping populations (see Shoemaker
et al., this issue). A large group of F2 progeny plants from A619 Ht2
X W64A, including many that were used here, were advanced to the F3 generation
by self-pollination. This will enable us to further test the current linkage
hypothesis in a more rigorous manner next year. We have also made the necessary
crosses to investigate the genetic relationship between Ht2 and
Ht3
in several inbred backgrounds.
Return to the MNL 66 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}