--M. Maddaloni, M. Albano, M. Motto and F. Salamini
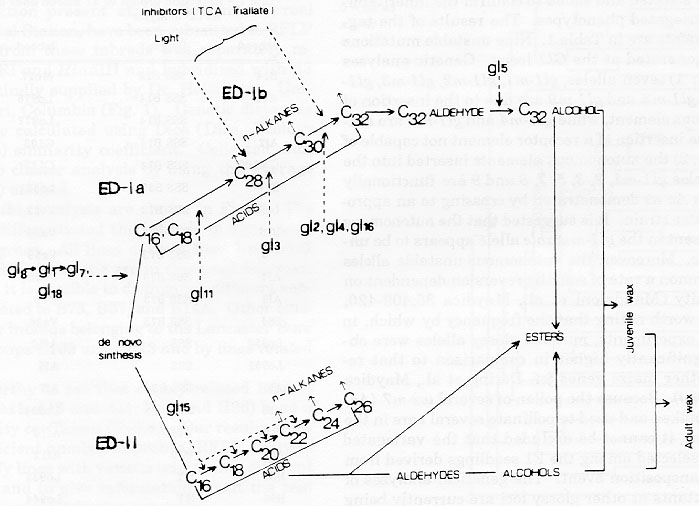
The first 5-6 leaves of maize seedlings are covered by a layer of wax which is characterized by a typical chemical composition (Bianchi et al., Maydica 30:179-198, 1985). Starting from the sixth leaf, the metabolic routes leading to the accumulation of the juvenile wax layer are clonally disactivated and only the adult waxes continue to be present, with a specific morphology and chemical composition. The biosynthetic pathway responsible for the accumulation of these organic compounds has been in part dissected by taking advantage of the glossy (gl) mutations (Fig. 1) (Bianchi et al.: quoted). However, our present knowledge at the biochemical level, of the mechanism o faction of the genes controlling wax deposition is unknown.
Because the molecular cloning of the glossy loci and the analysis of their structure and expression may shed light on the mechanism affecting waxy deposition, we have begun an extensive tagging program to generate unstable glossy alleles, at the most relevant loci affecting wax accumulation, with the aim to clone these genes. Homozygous plants from variegated wx-m7 kernels containing the Ac transposon at the Wx locus were crossed to several versions of the inbred line WF9, each containing stable double mutants at two different glossy loci. Variegated F1 seedlings, showing normal and glossy sectors on the leaves, were selected and selfed to confirm the inheritability of the variegated phenotypes. The results of the tagging experiments are in Table 1. Nine unstable mutations have been generated at the Gl1 locus. Genetic analyses showed that: 1) seven alleles, gl1-m1, gl1-m2, gl1-m3, gl1-m5, gl1-m7, gl1-m8 and gl1-m9 are due to the insertion of an autonomous element, while gl1-m4 and gl1-m6 are controlled by the insertion of a receptor element not capable of self-excision; 2) the autonomous elements inserted into the unstable alleles gl1-m1, 2, 3, 5, 7, 8 and 9 are functionally distinct from Ac as demonstrated by crossing to an appropriate Ac-tester strain. It is suggested that the autonomous element present in the gl1-mutable allele appears to be unrelated to Ac. Moreover, the autonomous unstable alleles share in common a rate of somatic reversion dependent on light intensity (Maddaloni et al., Maydica 35:409-420, 1990). It is worth noting that the frequency by which, in our tagging experiments, mutable glossy alleles were obtained is significantly higher in comparison to that reported for other maize genes (cf. Döring, H-P et al., Maydica 34:73-88, 1989). Because the pollen of several wx-m7 (Ac) plants was bulked and used to pollinate several ears in the original cross, it cannot be excluded that the variegated phenotypes selected among the F1 seedlings derived from the same transposition event. The genetical analyses of unstable mutants at other glossy loci are currently being addressed in our laboratory.
Figure 1. Biosynthetic scheme of maize surface wax assessed by studies on the effect of mutations, light, plant age and inhibitors on its composition.
Table 1. Summary of tagging experiments at relevant Glossy loci.
F2 confirmed mutable alleles
|
|
|
|
|
gl1 | gl8 | gl2 | gl5 | gl6 | gl3 |
| WF9gl1gl1gl8gl8 | Gl1Gl1Gl8Gl8 wx-m7 (Ac) |
|
|
|
|
|
|
|
|
| WF9gl2gl2gl8gl8 | Gl2Gl2Gl8Gl8 wx-m7 (Ac) |
|
|
|
|
|
|
|
|
| WF9gl5gl5gl8gl8 | Gl5Gl5Gl8Gl8 wx-m7 (Ac) |
|
|
||||||
| WF9gl2gl2gl6gl6 | Gl2Gl2Gl6Gl6 wx-m7 (Ac) |
|
|
||||||
| WF9gl3gl3gl8gl8 | Gl3Gl3Gl8Gl8 wx-m7 (Ac) |
|
|
||||||
| WF9gl2gl2gl3gl3 | Gl2Gl2Gl3Gl3 wx-m7 (Ac) |
|
|
Return to the MNL 65 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}