--M. M. Messmer, A. E. Melchinger, M. Lee and W. L. Woodman
We assayed maize inbred line B86, derived from the single cross B52xOh43, and its progenitors B52 and Oh43 with 187 mapped maize DNA clones in combination with three different restriction enzymes (EcoRI, EcoRV, and HindIII). The graphical genotype of B86 revealed by this RFLP analysis is shown in the previous note by Fähr et al.
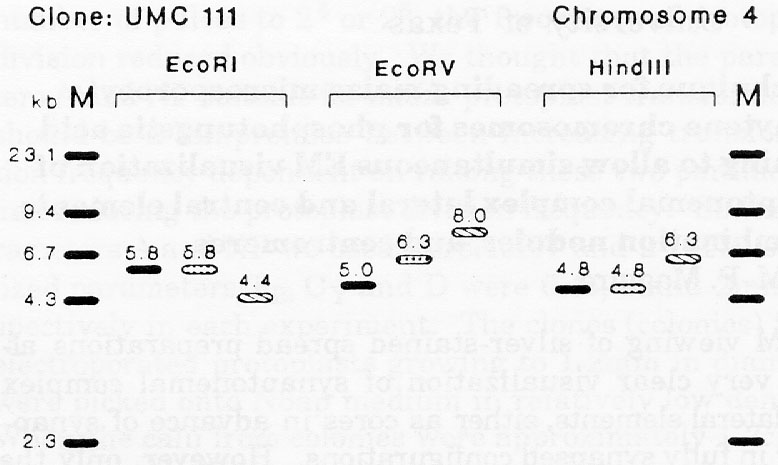
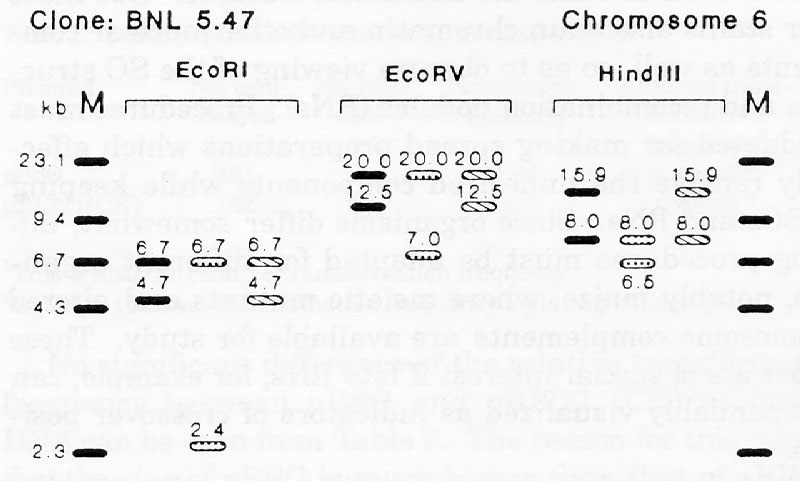
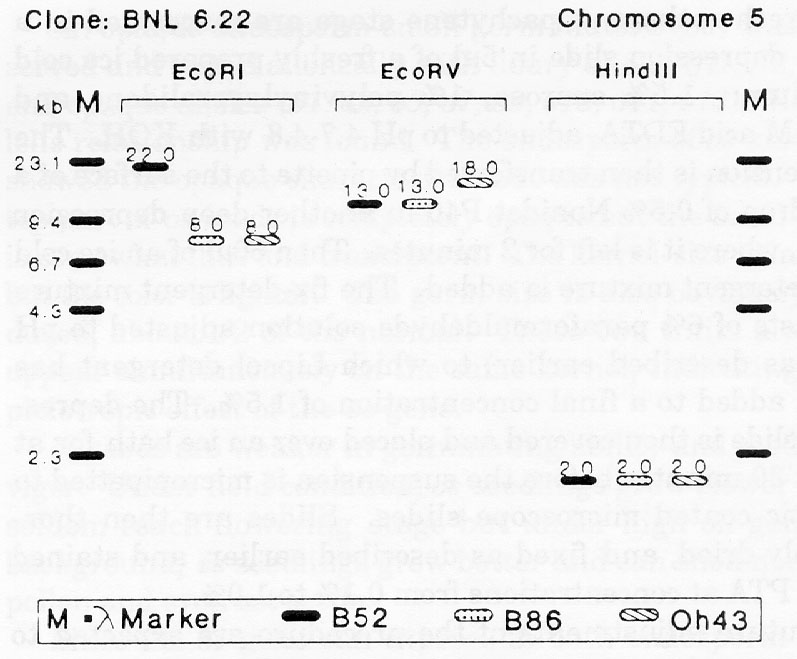
For clones UMC111, BNL5.47, and BNL6.22, previously located to chromosomes 4, 6, and 5, respectively, the RFLP banding patterns of B86 differed from those of its progenitors (Fig. 1). Explanations for the occurrence of new RFLP patterns in B86 could be residual heterozygosity or heterogeneity in the parental inbreds, mutation, or recombination during line development and maintenance. Pollen contamination or seed mixture are unlikely, because new RFLP variants in B86 occurred with a frequency of less than 2 percent. Also flanking markers were of parental types.
Based on the results of UMC111 with EcoRI and HindIII, B86 very likely inherited this genomic region from B52. The larger fragment of 6.3kb for B86 versus 5.0kb for B52 with EcoRV could be due to a point mutation that eliminated one restriction site for EcoRV in this region. This would agree with the observation of Fähr et al. that all RFLP patterns of B86 found on chromosome arm 4L were identical to those of B52. The RFLP pattern of B86 with UMC111 could also be explained by a crossover event within this region during line development, if B52 and Oh43 reveal class I RFLPs (R. B. Meagher et al., Genetics 120:809-818, 1988) with EcoRI and HindIII (i.e., the polymorphic fragments differ at only one end), but class II RFLP with EcoRV (i.e., the fragments differ at both ends).
The deviating RFLP patterns of B86 from those of B52 and Oh43 found with clone BNL5.47 and all three restriction enzymes could be caused by chromosomal mutation (deletion, insertion, or inversion) in this region in B86 that affects the bands of all three digests simultaneously. Another explanation for the different RFLP bands observed in B86 could be the presence of two RFLP variants in one of the parental lines. one of these variants was lost in the respective progenitor as a consequence of genetic drift, while the other was fixed in B86 during line development and maintenance.
The RFLP pattern of B86 for clone BNL6.22 was identical to Oh43 with EcoRI, but identical to B52 with EcoRV. This indicates a crossover event between the restriction sites of EcoRI and EcoRV in the early generations of line development, assuming class I RFLPs between B52 and Oh43. This matches with the graphical genotype of B86, where the RFLP pattern of the nearest clones, BNL10.06 and UMC67, originated from B52 and Oh43, respectively.
The deviations in the RFLP patterns of B86 from its progenitors for DNA clones UMC111 and BNL5.47 suggest that some instability at these RFLP loci could occur over several generations of line development and maintenance of B86 (1% of all probes examined). Further analyses with a larger set of materials, more DNA clones, and several restriction enzymes are needed for a more detailed investigation of (1) the molecular basis of these deviations in the RFLP patterns and (2) the frequency with which they occur.
Figure 1. RFLP banding patterns of inbred lines B52, B86, and Oh43 with
DNA clones UMC111,
BNL5.47,
and BNL6.22.
Return to the MNL 65 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}