Iowa State University
STUTTGART, GERMANY
University of Hohenheim
Graphical genotype for inbred line B86 determined by RFLPs
--S. Fähr, M. M. Messmer, A. H. Melchinger, M. Lee, and W. L. Woodman
As part of an extensive RFLP study of maize inbreds, we applied the concept of graphical genotypes (Young and Tanksley, Theor. Appl. Genet. 77:95-101, 1989) to maize inbred line B86. Our primary objective was to investigate the origin of the B86 genome from its parental inbreds B52 and Oh43.
Genomic DNA from all three inbreds (B52, Oh43, and B86) was digested separately with restriction enzymes EcoRI, EcoRV, and HindIII. RFLP analyses were performed with a total of 187 maize DNA probes (mainly genomic clones) kindly provided by four labs (B. Burr, Brookhaven National Laboratory; D. Grant, Pioneer Hi-Bred International Inc.; T. Helentjaris, Native Plants Inc.; D. Hoisington, University of Missouri-Columbia).
The DNA probes were positioned on the maize linkage map according to the most recently presented RFLP maps (MNL 64:154-163, 1990). The genotype of B86 for each marker was determined by comparing its RFLP patterns with those of B52 and Oh43.
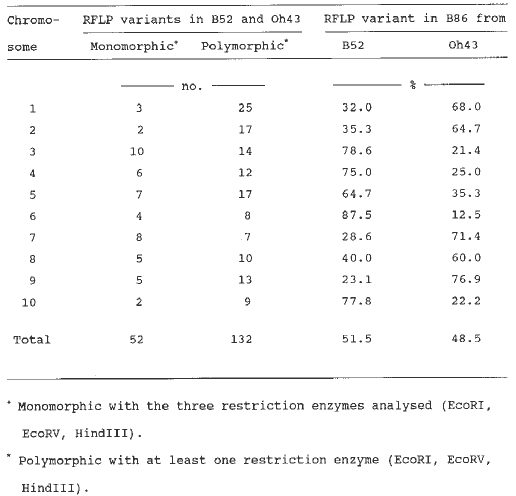
Three probes yielded deviating RFLP patterns for B86 not present in the parental inbreds (details are given in the following note of this MNL by Messmer et al.). Altogether, 132 (71.7%) out of the remaining 184 DNA probes revealed polymorphic RFLP patterns in B52 and Oh43 and only these are subsequently considered. RFLP variants in B86 at loci mapping to chromosomes 3, 4, 5, 6, and 10 originated mostly from B52 whereas those mapping to chromosomes 1, 2, 7, 8, and 9 originated mostly from Oh43 (Table 1). Averaged across the whole genome, B86 inherited 51.5% of the RFLP variants from B52 and 48.5% from Oh43.
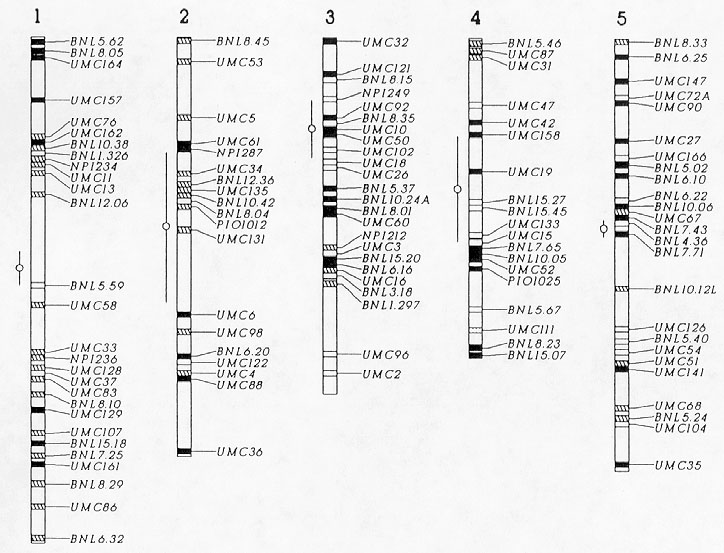
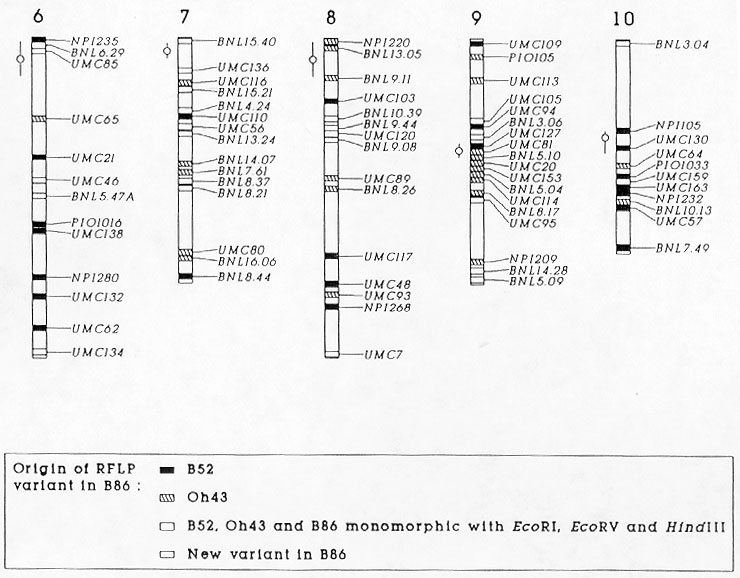
Figure 1 shows a graphical representation of the origin of the RFLP variants detected in B86. Chromosomal regions, in which several adjacent markers had the same origin of RFLP variations, were probably transmitted from the respective parent without effective recombination events. Examples include chromosome arms 8S and 9L, probably inherited intact from Oh43, and chromosome arm 3S from B52. In general, the graphical genotype provides information about the number of effective recombination events that accumulated during the selfing process and in the development of B86. However, our results must be interpreted with caution because valid conclusions about recombination events could only be drawn based on an RFLP linkage map especially constructed for cross B52xOh43. Despite this limitation it seems safe to assume that a recombination event occurred on the short arm of chromosome 4, because the RFLP variants at the three distal markers came from Oh43, whereas the remaining part of the chromosome was very likely inherited from B52.
B86 is a selection from a breeding program conducted by W. A. Russell and W. D. Guthrie at Iowa State. The aim of selection was to combine leaf-feeding resistance (1st brood resistance) and sheath and collar-feeding resistance (2nd brood resistance) to the European corn borer (ECB) Ostrinia nubilalis Hübner into a single genotype. Leaf-feeding resistance was available in Oh43, which is moderately resistant to 1st brood and has good combining ability for grain yield but is highly susceptible to 2nd brood (Russell et al., Crop Sci. 14:725-727, 1974). B52 is highly resistant to 2nd brood and also shows intermediate resistance to 1st brood but has poor yield performance (Russell and Guthrie,, WD Crop Sci. 19:565, 1979). B86 was the best inbred developed from 200 F3 lines of the cross B52xOh43 by selection and selfing according to the ear-to-row method from the F3 to F7 generations, using artificial infestations with ECB egg masses in each generation (Russell et al., 1974, see above; Guthrie et al., J. Econ. Entomol. 78:93-95, 1985). The main selection criteria were resistance to both generations of ECB and, within highly resistant progenies, recovery of the Oh43 phenotype (W. A. Russell, pers. comm.). B86 proved to be highly resistant to 1st brood and nearly as resistant as B52 to 2nd brood of ECB.
The estimated number of genes conferring resistance to 2nd brood of ECB in B52 is seven (Onukogu et al., J. Econ. Entomol. 71:1-4, 1978). Russell et al. (1974) assumed three loci conditioning 1st brood resistance in Oh43 and a different locus in B52, but absence of genes for 2nd brood resistance in Oh43. These authors also reported that the genes conferring the two types of resistance are not identical. Based on this information, it seems reasonable to assume that approximately eight loci from B52 and three loci from Oh43 were under intense selection pressure during the development of B86. Thus, B86 is expected to have a higher proportion of RFLP variants from B52 than from Oh43, which deviates from the observed ratio of approximately 1:1 (Table 1).
So far, no genes conferring resistance against 1st or 2nd brood of ECB have been reported for chromosomes 7, 9, and 10. With regard to these chromosomes, 41.4% of the polymorphic variants observed in B86 were derived from B52. This suggests that parts of the B86 genome, having no effect on ECB resistance, were mainly inherited from Oh43 due to selection for recovery of the agronomically desirable Oh43 phenotype. B86 probably inherited four genes conditioning 1st brood resistance to ECB (three from Oh43 and one from B52), but not all seven genes from B52 conditioning 2nd brood resistance to ECB, because it is slightly less resistant to 2nd brood than B52. In fact, a higher proportion (54.4%) of RFLP variants in B86 on chromosomes 1, 2, 3, 4, 5, 6 and 8, which are supposed to carry resistance genes to 1st and 2nd brood, were derived from B52.
In addition to selection for genes conditioning the expression of different traits from each parent, genetic drift of genomic regions with neutral effect on the selection criteria would result in random fixation of RFLP variants from each parent. Thus, random genetic drift might be another explanation for the observed 1:1 ratio of RFLP variants of B52 and Oh43 found for the molecular genotype of B86 in this study.
Table 1. Number of DNA probes, yielding monomorphic and polymorphic RFLP variants for maize inbreds B52 and Oh43, associated with each of the 10 maize chromosomes. For polymorphic RFLP variants, the last two columns show the origin of the RFLP variants in inbred B86.
Return to the MNL 65 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}