--Donald S. Robertson and Philip S. Stinard
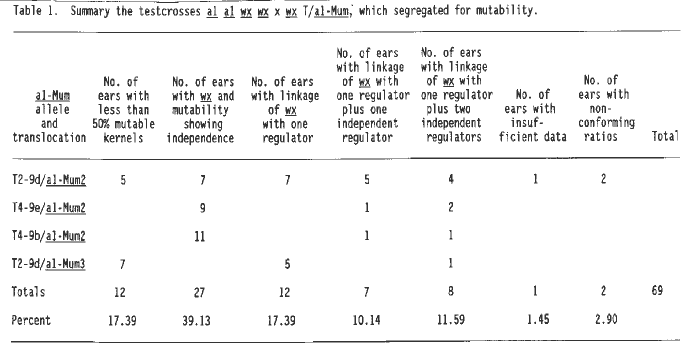
In last year's Newsletter (MNL 64:13, 1990) we presented preliminary data indicating that a regulator of the somatic mutability of the Mutator-induced a1-Mum3 allele was located on chromosome two. These data were based on only a few crosses involving wx T2-9d (2L.83, 9L.27). Additional kernels from the F1 ears (wx T2-9d/a1-Mum3) that had indicated linkage were planted in 1990 and the resulting plants crossed to a1 wx stocks. In 1989, a new set of crosses was made to the waxy marked translocation series using plants grown from mutable kernels of outcross ears segregating for one regulator. These involved a1-Mum2 and a1-Mum3 stocks, for which we had not previously obtained any positive tests for linkage. In the winter nursery of 1989-90, the F1's of this new set of crosses were testcrossed to a1 wx stocks. The resulting ears were screened for indications of linkage of a regulator with wx. Kernels from ears yielding F1 plants that showed linkage in the winter nursery were planted in large numbers (30 to 100 kernels, depending upon the F1) in the summer of 1990 and the resulting plants were testcrossed to a1 wx testers.
The results are summarized in Table 1. Although all the plants used for crossing to the translocation series grew from mutable kernels of ears segregating in a 1:1 ratio for mutable and stable kernels, some of the progeny, when tested, gave ratios that indicated there had been replication of the regulator resulting in plants with two or three regulators. The outcrosses of these plants to the translocation series were also grown to determine if linkage of a regulator with more than one translocation could be demonstrated.
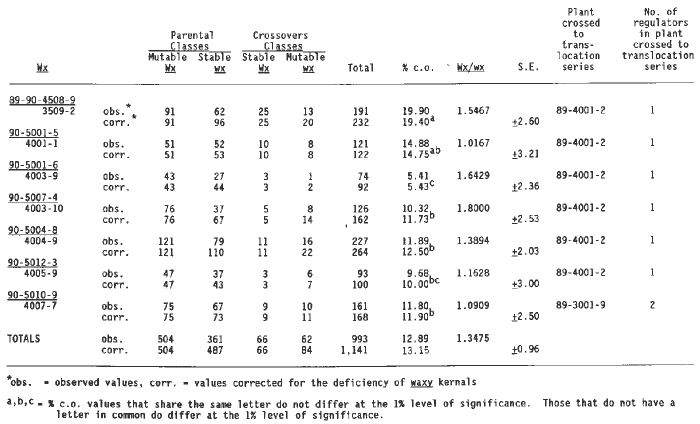
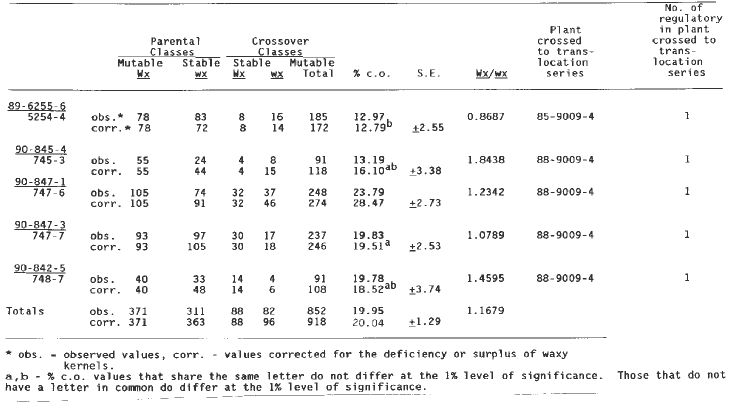
The crosses of a-Mum2 with only one element linked to waxy (Table 2) showed some variation in the amount of crossing over; from a low of 5.43% to a high of 19.40%. There seem to be three levels of recombination (i.e., 5-10%, 10-15%, 15-20%) based on a statistical analysis of the differences between the different recombination values. If these are real differences in recombination, as the statistics indicate, these data would suggest that the regulator has transposed from one location on chromosome two to another. Based on the raw data, the average crossing over for all seven crosses is 12.89%. However, in most crosses there is a deficiency of waxy kernels. The reduced transmission of the wx allele by heterozygous Wx wx plants has been reported in the literature. The �corrected totals� in this and other tables have been calculated by increasing the waxy classes to make them equal to the starchy classes. The added waxy values were distributed among the mutable and stable classes in the same proportion they were observed in the raw data. With this correction in Table 2, the average percentage crossing over is increased from 12.89% to 13.15%. Plants grown from kernels of two different 1:1 ears (i.e., 89-4001-2, 89-3001-9) were crossed to wx T2-9d. The crossover value for the one regulator outcross of 89-3001-9 is very close to some of the crossover values observed for the outcrosses of 89-4001-2. These 1:1 ears may have carried the same regulator at the identical locus as the result of descent from a common progenitor (i.e., the original a1-Mum2 mutation event). Alternatively, their common values may be just coincidental.
Plant number 89-3001-9 showed evidence of carrying two regulators. The testcross with wx T2-9d (Table 2) indicated the presence of only one regulator. Outcrosses of this same plant to other translocations gave results consistent with the presence of 2 or 3 regulators in different F1's (Tables 3 and 4), thus demonstrating that the number of regulators can either increase or decrease with a single generation of outcrossing.
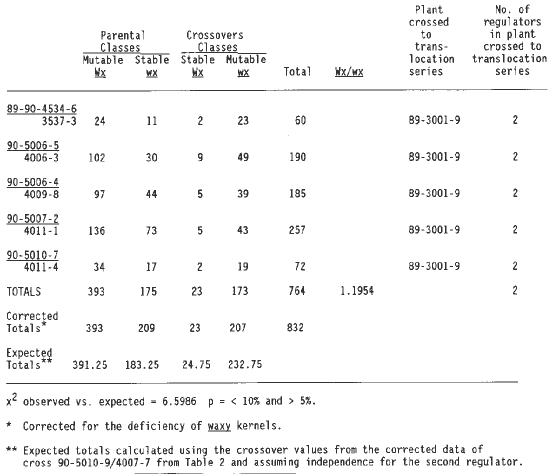
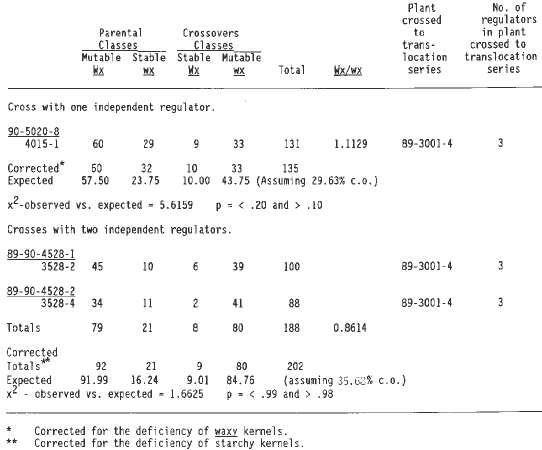
Table 3 summarizes the data from crosses with a regulator linked to wx T2-9d plus a second independent regulator. The observed and expected results are in good agreement. The expected results were calculated by using the corrected linkage value of the last cross of Table 2, because this cross and all the crosses in Table 3 shared the same 1:1 parent plant (89-3001-9). Thus, it is likely that the regulator linked to wx T2-9d is at the same position in both the F1 with one linked regulator and the F1's with a second independent regulator.
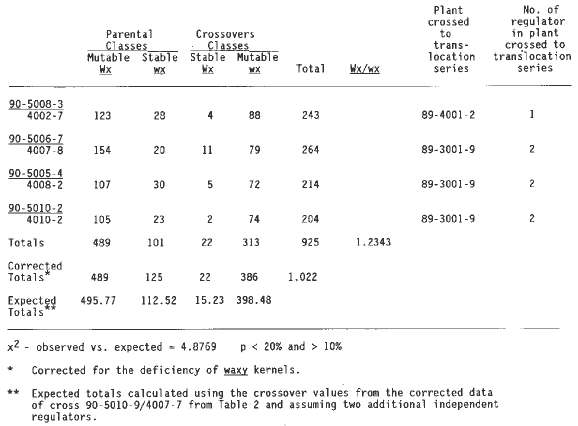
Table 4 summarizes the data from crosses with a regulator linked to wx T2-9d and two additional independent regulators. Again the corrected linkage value for the last cross in Table 2 was used to calculate expected values, because all but one cross in Table 4 involved plant 89-3001-9. The data do not differ significantly from the expected values. The first cross of Table 4 came from an a1-Mum2 plant (89-4001-2) that had only one regulator and yet now three are observed, one linked to wx T2-9d and two others at independent locations. Plant 89-3001-9 contributed to testcrosses in Tables 2, 3 and 4. This plant carried two independent regulators, one of which is shown to be linked to wx T2-9d (Table 2). Because the second regulator is not linked to wx one third of the plants segregating for somatic mutability would be expected to receive only the linked regulator, one third only the nonlinked regulator and one third both regulators. One of the outcrosses carried only the linked regulator (Table 2), and seven carried the unlinked regulator (the 7 crosses shown in line one column three of Table 1), and eight had both regulators (Tables 3 and 4).
Three crosses involving wx T4-9e (4S.53, 9L.26) and a1-Mum2 gave plants with one element linked to this translocation plus one independent regulator (one plant) or two independent regulators (two plants, Table 5). For both of these situations, the observed values do not differ significantly from the predicted values estimated by using the frequency of the starchy stable classes (0.0741 and 0.0446 respectively) to calculate the expected frequencies. Using these values gives estimates of wx to regulator distance of 29.64 and 35.68 respectively. All of the crosses of Table 5 are derived from the same a1-Mum2 plant, which had 3 regulators. Thus, it is possible that the same regulator located at the same position relative to the translocation breakpoint is present in the plants with one and two independent regulators. The difference between the two crossover values (6.04%), is not statistically significant.
The a1-Mum2 plant crossed to wx T4-9e (89-3001-4, Table 5) was a sibling plant to the one that yielded one of the crosses that had one regulator linked to wx T2-9d (89-3001-9, Table 2), all of the crosses with one regulator linked to wx T2-9d plus an independent regulator (Table 3), and three out of four crosses with one regulator linked to wx T2-9d plus two independent regulators (Table 4). It is conceivable that one of the independent regulators of Table 5 is the regulator linked to wx T2-9d of Tables 2, 3 and 4, although this can not be proved from the existing data.
Two crosses of wx T4-9b/a1-Mum2 gave results consistent with a regulator linked to wx T4-9b (4L.90, 9L.29) and one or two additional independent regulators (Table 6). The crossover value and the chi-square test for the cross with two regulators is not reliable because one class (starchy, stable) had only one kernel and because this was the class used to estimate the amount of crossing over. However, all classes show close agreement between observed and expected values. Thus there is a strong indication of linkage of a regulator with the wx T4-9b breakpoint, with a crossover value that probably is not greatly different from the estimated one. There is little likelihood that the linked regulators in the one independent and the two independent regulator crosses are at the same positions because of the large difference in recombination values
(Table 6). The a1-Mum2 plant outcrossed to wx T4-9b (89-3001-1T, Table 6) was a sibling plant to the one crossed to wx T4-9e (89-3001-4, Table 5) and also a sibling to one of those crossed to wx T2-9b (89-3001-9, Tables 2, 3 and 4). Again it is possible, but by no means certain, that an independent regulator in the wx T4-9b multiple regulator crosses could be the one mapped in the wx T2-9d crosses.
Evidence for linkage of a regulator to wx T2-9d was also found in a 1:1 a1-Mum3 test (Table 7). The crossing over for the total population was 19.95 percent (corrected 20.04 percent), which is significantly higher than that observed for the a1-Mum2 linkage with the same translocation (13.15%, Table 2). Some of the individual a1-Mum3 crossover frequencies, however, do not differ significantly from those observed in the a1-Mum2 crosses. Values of between 12 and 20 percent are found in both sets of testcrosses. (The Table 2 crossover percentages of 14.75, 12.50, 11.73, 11.90 and 10.00 do not differ significantly at the one percent level from those of 19.51, 18.52, 16.10 and 12.79 of Table 7. The crossover percentage of 19.40 in Table 2 is not significantly different from those of 19.51, 18.52 and 16.10 of Table 7.) One a1-Mum2 cross has a crossover value considerably less (5.43 percent) than any found for a1-Mum3 crosses and one a1-Mum3 cross has a value of 28.47 percent, which is considerably higher than any found for a1-Mum2. The a1-Mum2 and a1-Mum3 alleles were produced in the same isolation plot, in which purple aleurone Mutator plants were used as male parents. It is possible that these two mutant alleles arose from a sector in the tassel of a purple Mutator plant and thus might have had some regulators in common. Analyses of the original isolates of these mutants in their immediate progenies and in later generations indicated that multiple regulators of somatic mutability were present in the genome of these mutants when they were first isolated. Even if these are not alleles derived from a single mutation event but arose independently they nonetheless were induced in a common purple aleurone Mutator population, and thus progeny plants carrying the independent mutations could have many regulators in common. Therefore, it is possible that lines carrying the different a1-Mum alleles have a common regulator on the long arm of chromosome two.
One test cross of a wx T2-9d a1-Mum3 F1 segregated for a regulator linked to the translocation plus two independent regulators (Table 8). The chi-square in this instance is not reliable because only one starchy stable kernel was observed. However, the expected frequencies of all classes are in close agreement with the observed values, suggesting that this cross indeed involves two independent regulators.
These studies are illustrative of how readily the number of regulators can change. Plant 89-3001-9 (a1-Mum2), with two regulators, was outcrossed to the translocation series, followed by testcrossing the next generation. In three of the progeny of the crosses of this plant to wx T2-9d that showed linkage, the number of regulators had increased from the two found in the parent to 3 in the testcrosses (Table 4). One sibling plant of 89-3001-9, 89-3001-1T, also went from two regulators in the parent to three in the testcross (Table 6). The a1-Mum3 plant which was crossed to wx T2-9d, 88-9009-4, had only one regulator, and in most testcrosses of the F1 only one was retained. However, in one cross two additional regulators were found (Table 8).
Conclusions. 1) Ears that have 1:1 or other ratios of mutable:stable kernels, indicative of the presence of one or more regulators of somatic mutability, have been shown to be carrying regulators that can be mapped to chromosome locations using standard linkage tests. In the stocks tested, regulators have been located to the long arm of chromosome 2 and to the long and short arms of chromosome 4.
2) In different sibling plants from an outcross of a single plant to wx T2-9d, several different crossover values are observed, suggesting that the regulator on chromosome two may be transposing to different positions on the chromosome.
3) Regulators of somatic activity of Mutator-induced mutants can both increase and decrease in number from one generation to the next. In these studies, the increase in number is probably the result of the transposition of the regulator to a locus independent of its original site. The decrease in number is undoubtedly the result of the independent assortment of unlinked regulators.
Table 1. Summary of the testcrosses a1 a1 wx wx x T/a1-Mum, which segregated for mutability.
Table 2. Testcrosses of wx T2-9d/a1Mum2 with a1 a1 wx wx showing linkage of wx with one regulator element.
Table 3. Testcrosses of wx T2-9d/a1Mum2 with a1 a1 wx wx showing linkage of wx with one regulator element plus a second independent regulator.
Table 4. Testcrosses of wx T2-9d/a1Mum2 with a1 a1 wx wx showing linkage of wx with one regulator element plus 2 independent regulators.
Table 5. Testcrosses of wx T2-9e/a1Mum2 with a1 a1 wx wx showing linkage of wx with one regulator element plus one or two independent regulators.
Table 6. Testcrosses of wx T2-9b/a1Mum2 with a1 a1 wx wx showing linkage of wx with one regulator element plus one or two independent regulators.
Table 7. Testcrosses of wx T2-9d/a1Mum3 with a1 a1 wx wx showing linkage of wx with one regulator element.
Table
8. Results of a testcross of wx T2-9d/a1Mum3 with
a1 a1 wx wx showing linkage of wx
with one regulator plus two independent regulators.
Return to the MNL 65 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}