Northrup King Co.
Near-isogenic line localization of MDMV resistance to chromosome 6S
--Ed Weck, Diana Beckman, Doug Mead, Christi Bredenkamp and Mary Trainor
A single major gene for MDMV resistance (Mdm1) was mapped to chromosome 6S between probes UMC85 and BNL6.29 (McMullen and Louie, Mol. Plant-Microbe Interactions 2:309, 1989) in two backcross populations using the inbred Pa405 as a source of resistance because of its symptomless response to MDMV strains A and B. Near-isogenic lines are a genetic resource for the integration of conventional and molecular marker linkage maps (G. J. Muehlbauer, et al., Crop Sci. 28:729, 1988). We have confirmed the chromosome 6S location for MDMV resistance using two independently created near-isogenic lines. Selections for MDMV resistance with 6S markers confirm the location of MDMV resistance.
Table 1. Lines analyzed and their level of MDMV resistance.
| Line | % MDMV Resistant |
| Pa405 | 100% |
| NKA | 0% |
| NKA-MDM (Mdm1 from Pa405) | 100% |
| NKB | 0% |
| NKB-MDM (Mdm1 from Pa405) | 66% |
| NKC = (NKC*8/Pa405) 7x | 86% |
| NKD = (NKD*9/Pa405) 7x | 58% |
| NKE = (NKE*8/Pa405) 6x | 21% |
Two independently created MDMV resistant lines (NKA-MDM,NKB-MDM), the near-isogenic non-resistant lines (NKA,NKB) and the source of MDMV resistance (Pa405) were analyzed with 139 RFLP probes distributed over all corn chromosomes, see Figure 1. Two probes, NPI7 and UMC85 on chromosome 6S, showed a common pattern for the MDMV resistant lines: NKA-MDM; NKB-MDM; and Pa405. The presence of this chromosome 6S region in two independent MDMV resistance conversions and in the backcross populations of McMullen and Louie (Mol. Plant-Microbe Interactions 2:309, 1989) indicates that this is an important region in MDMV resistance.
Other regions from Pa405 appeared in the MDMV resistance conversions, NKA-MDM and NKB-MDM. Two probes represented by the letter B, showed an identical banding pattern for NKB-MDM and Pa405 and one probe, represented by the letter A, showed an identical banding pattern for NKA-MDM and Pa405. Letters A and B indi-cate regions of the Pa405 genome that were transferred to MDMV conversions NKB-MDM or NKA-MDM respectively. The additional region in NKA-MDM or the two additional regions in NKB-MDM could result in NKA-MDM being 100% resistant while NKB-MDM is only 66% resistant.
The location of Mdm1 on chromosome 6S was used to guide MDMV resistance selection in lines segregating for
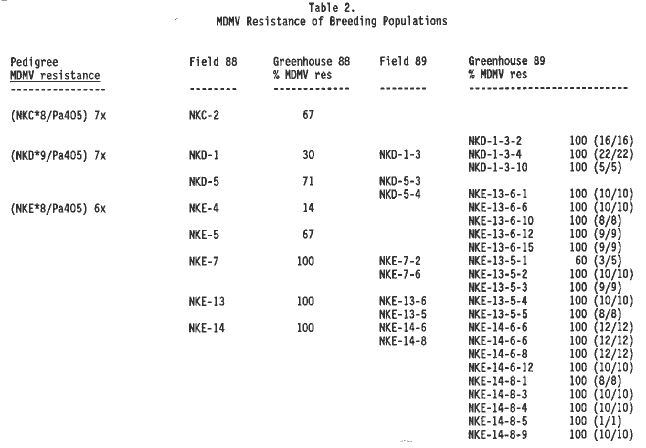
MDMV resistance which had not yielded to breeders attempts to fix the resistance. In the summer of 1988 three lines were planted: (NKC*8/Pa405)7x, (NKD*9/Pa405) 7x and (NKE*8/Pa405) 7x. Table 2 shows the selections planted in the field and greenhouse and the percentage of plants resistant to MDMV. Pollinations from these populations were analyzed for MDMV resistance in the winter of 1988 in the greenhouse. Selections were planted in the summer of 1989, analyzed with chromosome 6S probes, and tested for MDMV resistance in the greenhouse in the winter of 1989. The winter 1989 MDMV viral greenhouse screen showed 99% (204/206) of the individuals to be MDMV resistant following selection with 6S probes NPI7 and UMC85 in the summer of 1989. This indicates that these two 6S probes are strongly correlated with MDMV resistance when using Pa405 as a source.
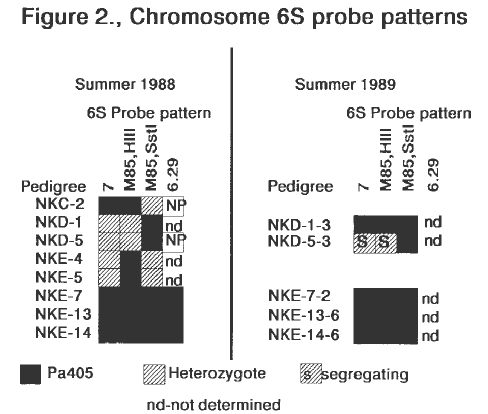
Probe patterns for 1988 and 1989 field grown plants are summarized in Figure 2. MDMV resistance data for these plants is shown in Table 2. Genomic DNA was digested with enzymes SstI and HindIII in an attempt to ex-plain the different patterns observed with probe UMC85 relative to the level of MDMV resistance. Plants which were 100% resistant had the Pa405 pattern for both enzymes. Probe BNL6.29 was non-polymorphic for NKD-1, NKD-5 and NKC-2.
We have shown that near-isogenic lines can be used in conjunction with RFLPs to localize traits to a marker interval. Two independent near-isogenic conversions may be sufficient to localize a trait of interest and to eliminate regions from the nonrecurrent parent genomes unrelated to that trait. An advantage of the use of near-isogenic lines is that fewer lines are required for molecular marker analysis. A disadvantage is the number of generations required to construct near-isogenic lines by backcrossing. Near-isogenic lines are a powerful mapping tool in crops such as tomato and soybean where a large number of near-isogenic lines are available.
Figure 1. Probe distribution.
Figure 2. Chromosome 6S probe patterns.
Table
2. MDMV Resistance of Breeding Populations.
Return to the MNL 65 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}