University of Wisconsin
Preliminary results from the molecular analysis of intragenic recombination at the waxy locus
--Ron J. Okagaki
The waxy locus has been a focus for research on intragenic recombination in higher plants for almost 40 years. Most of the information on intragenic recombination is limited to genetic phenomenology, however this situation should change with the molecular characterization of these events. Ultimately we hope to come to an understanding of the rules governing the behavior of mutant heteroallelic combinations. In this report, data from Nelson's genetic studies on intragenic recombination at the waxy locus (MNL 50:109, 1976) is combined with molecular studies on the locus from the Wessler lab (PNAS 82:4177, 1985; PNAS 87:8731, 1990; and unpublished data) and other sources (Klosgen et al., MGG 203:237, 1986). This extensive body of data is available at the waxy locus first because of the large numbers of mutant alleles that have been recovered and second because the waxy phenotype can be determined by staining pollen with iodine. It was this ability to screen thousands of wx pollen grains for rare revertant Wx pollen grains that has made these studies possible.
Two aspects of intragenic recombinations are discussed here, the relationship between intragenic recombination frequency and the physical distance separating mutant lesions and second the effect of the size of mutant lesions on the ability of a mutant allele to recombine. Lastly, the outline of an ongoing project designed to study gene conversion and estimated the size of conversion tracts is presented.
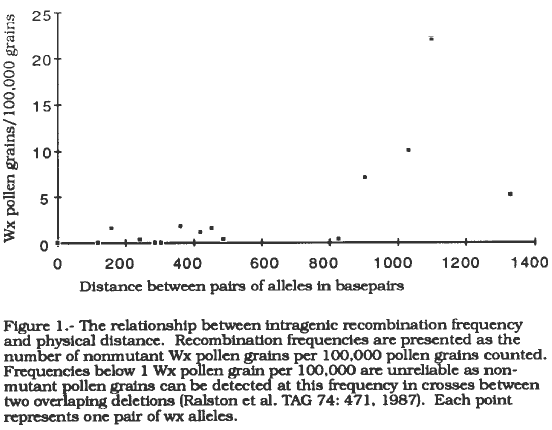
A simplistic model of intragenic recombination would assume that the probability of intragenic recombination occurring between two sites is largely determined by the distance between the two sites. According to this model as the physical distance between two mutant lesions increases the frequency of recovering nonmutant alleles also increases. Figure 1 presents a graph of this relationship in waxy alleles which shows a weak association between increasing recombination frequency and increased physical distance. However, distance alone is not a strong predictor of recombination frequency between two alleles suggesting that additional factors influence intragenic recombination.
One possible explanation for the scatter in the data is the size of the lesion. Work at the bronze-1 locus has shown that Ds insertions compress the genetic fine structure map (Dooner, Genetics 113:1021, 1986), and to date only insertions and deletions have been placed on the waxy physical map. Insertions in the waxy locus range in size from approximately 0.15kb for wx-B2 to over 5kb for wx-Stonor and wx-M, and deletions range from 30bp for wx-C and wx-BL2 (note, these alleles have identical breakpoints) to wx-C34 which deletes the entire locus. This approach takes into account both the distance between lesions and the size of lesions, and predicts that increasing the size of the lesion will decrease the rate of recombination. Data at the waxy locus do not allow the testing of this possibility. The difficulty lies with the confounding of the variables distance and size. It is likely that the size of lesions will affect intragenic recombination as has been found at the bronze-1 locus, but demonstration of this will require additional data. One observation can be made for alleles with large insertions, such as wx-M and wx-Stonor which are larger than the waxy locus itself; these insertion alleles are able to recombine with other alleles within the waxy locus. Insertions and deletions can inhibit intragenic recombination, but this suppression may be limited to short distances.
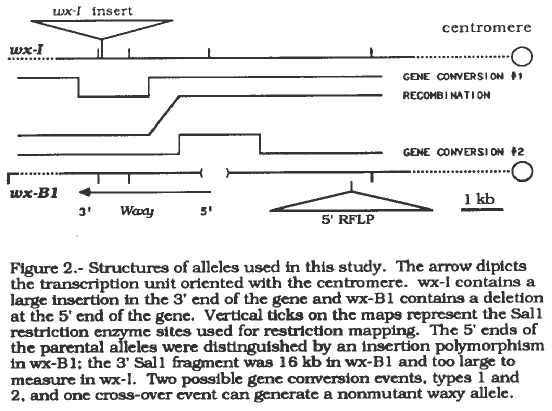
In an attempt to learn more about intragenic recombination a project to study gene conversion and estimate the size of conversion tracts was initiated. The molecular descriptions of the three mutant waxy alleles used in this study, wx-C34, wx-B1, and wx-I, have previously been published (PNAS 82:4177, 1985). wx-C34 is a deletion of the entire waxy locus, and crossing the Wx* alleles produced by intragenic recombination with wx-C34 permits the analysis of Wx* alleles without the complication of other waxy sequences in the genome. wx-B1 and wx-I were chosen because Southern blot analysis can distinguish the lesions causing the mutations as well as the 5' and 3' ends of the alleles (Figure 2).
Homozygous wx-B1 and wx-I plants were crossed to produce the heteroallelic mutant; these plants were crossed by wx-C34 and nonmutant Wx* plants. Wx kernels arising from pollination by stray Wx pollen were identified at this stage. Contaminants will contain a nonmutant Wx allele and either a wx-I or wx-B1 allele instead of a Wx* allele over the deletion of the waxy locus.
Wx* alleles arising from gene conversion versus recombination were also distinguished by Southern blot analysis. Recombination generates Wx* alleles with 5' flanking RFLPs from wx-I and 3' RFLPs from wxwx-B1 (Figure 2). Wx* alleles created through gene conversion should resemble one of the parental alleles unless the conversion tract is very large. To date four Wx* alleles have been partially characterized. One allele arose through recombination, and three alleles have the flanking RFLPs of wx-I and apparently arose by gene conversion of the type 1 class (Figure 2). one putative Wx* allele was determined to be a contaminant, and 35 additional Wx* alleles have been isolated and await analysis.
Figure 1. The relationship between intragenic recombination frequency and physical distance. Recombination frequencies are presented as the number of nonmutant Wx pollen grains per 100,000 pollen grains counted. Frequencies below 1 Wx pollen grain per 100,000 are unreliable as non-mutant pollen grains can be detected at this frequency in crosses between two overlaping deletions (Ralston et al TAG 74: 471, 1987). Each point represents on pair of wx alleles.
Figure
2. Structures of alleles used in this study. The arrow
dipicts the transcription unit oriented with the centromere. wx-1
contains a large insertion in the 3' end of the gene and wx-B1 contains
a deletion at the 5' end of the gene. Vertical ticks on the maps
represent the Sal1 restriction enzyme sites used for restriction mapping.
The 5' ends of the parental alleles were distinguished by an insertion
polymorphism in wx-B1; the 3' Sal1 fragment was 16 kb in wx-B1 and too
large to measure in wx-1. Two possible gene conversion events, types
1 and 2, and one cross-over event can generate a nonmutant waxy allele.
Return to the MNL 65 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}