--Víctor Rául Corcuera
At the latitude in which most cultivated maize is concentrated in Argentina (35º southern latitude), normally all the teosinte species express a strong photoperiod response, flowering during autumn (April-May). Thus, it can be said that they show a long evolutive cycle. Under these environmental conditions, annual teosintes like diploperennial teosintes have an evolutive cycle measured in number of days from plant emergence up to tasselling that is 3 times longer than cultivated maize (190 days against 60 days for maize). It can also be added that annual teosintes must accumulate 560 C to tassel, whilst maize only needs approximately 250 C.
Evolutive cycle traits were measured in 7 different populations to obtain the basic information for future projects in which annual teosinte germplasm could be used for maize genetic improvement. The 7 populations studied are: maize inbred line OU (P1), Balsas teosinte (P2), Guatemala teosinte (P3), and the F1 and F2 progenies derived from crossing maize with each one of these teosinte species.
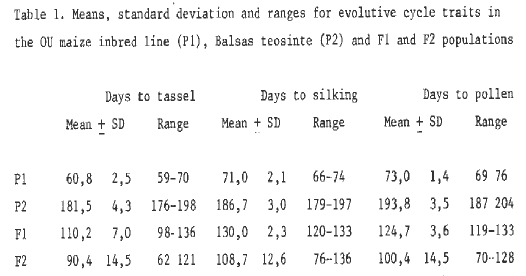
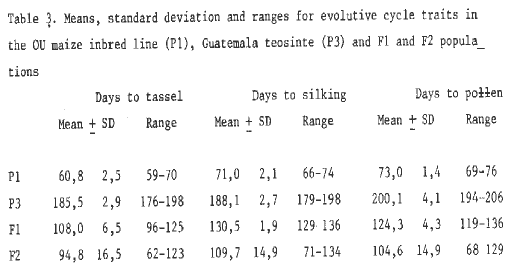
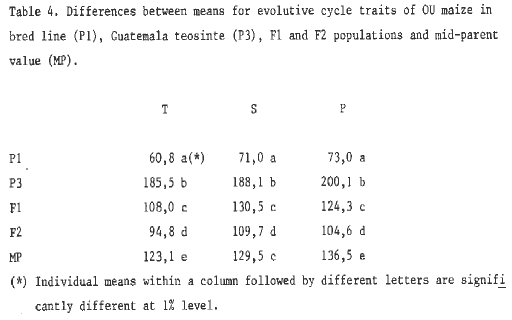
On the basis of individual plants, cycle traits measured were: 1) days to tassel (T), 2) days to silking (S) and 3) days to pollen (P). The results obtained are summarized in Tables 1 to 4. It can easily be noted that in both crossings the average values both for the F1 and F2 progenies are smaller than the mid-parent value (MP). At the same time, the amplitude of variation for each trait considered is wide enough to reach their parent extreme values (see Tables 1 and 3).
The results obtained point out that cycle traits are quantitatively inherited: a short cycle (maize) is dominant, though in a medium degree, over a long cycle (teosinte). This fact indicates that maize-Balsas and maize-Guatemala teosinte hybrids behave in a similar way to diploperennial teosinte-maize hybrids (see MNL 62:77, 1988).
In addition, the evolutive cycle traits measured are closely entailed among themselves and their hereditary behaviour is very similar. Otherwise, as in previous publications (see MNL 62:77, 1988), these hybrids are protandrous.
The results obtained demonstrate that it would be relatively easy to select short evolutive cycle individuals derived from these crossings, though this is not a surprising fact, as previous investigations we have done already demonstrate it. Nevertheless, it is really very important and advantageous that individuals derived from these non-traditional crosses and expressing a short cycle duration can be chosen amongst the whole, as they may be grown in the field and mature under normal environmental conditions.
Table 1. Means, standard deviation and ranges for evolutive cylce traits in the OU maize inbred line (P1), Balsas teosinte (P2) and F1 and F2 populations.
Table 2. Differences betweens means for evolutive cylce traits in the OU maize inbred line (P1), Balsas teosinte (P2), F1 and F2 populations and mid-parent value (MP).
Table 3. Means, standard deviation and ranges for evolutive cylce traits in the OU maize inbred line (P1), Guatemala teosinte (P3) and F1 and F2 populations.
Table
4. Differences betweens means for evolutive cylce traits in the
OU maize inbred line (P1), Guatemala teosinte (P3), F1 and F2 populations
and mid-parent value (MP).
Return to the MNL 65 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}