Krasnodar Research Inst. Agric.
Genetic control of maize variability
--A. S. Mashnenkov
Actual progress in evolution and breeding is attributed to different types of genetic variability: mutation, recombination, combination, introgression. The interaction of these sources of variability controlled by inheritance is not known to the author.
At first we managed to reveal a curvilinear association between maize inbred sensitivity to N-nitroso-N-methylurea in M1 and their mutability (MNL 59:104). Some insensitive and sensitive low-mutable maize inbreds proved similar to Uvr+RecA+ and Uvr-RecA- of E. coli strains respectively, and those of moderate sensitivity high-mutable inbreds to strain Uvr-RecA+. We supposed the existence of an ancient system of genome maintenance providing distinction and removal of any distortions in genetic substance (Genome 30:159, 1988). Proceeding from this hypothesis the following considerations may be drawn:
1. In case of distant hybridization with low-mutable lines insensitive to mutagen effects hybrid kernels would not develop. In high-mutable lines the maximum frequency of hybrid seed set is expected.
2. In low-mutable sensitive inbreds elimination of alien genetic stock produced through distant hybridization or transformation rarely occurs.
3. In supersensitive inbreds recombination processes are partially or completely suppressed.
Our experience in crossing maize with Tripsacum dactyloides (2n=72) mainly proves consequence 1 of the general hypothesis.
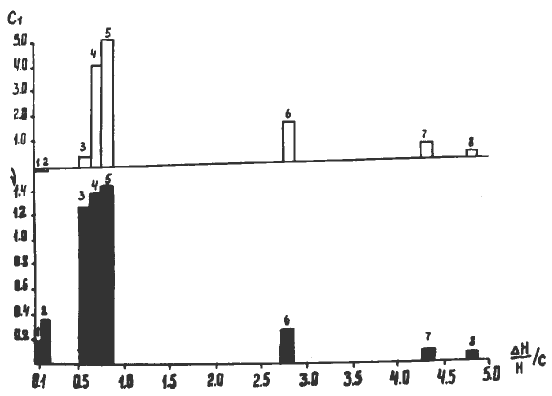
Some considerable deviation from the theoretically expected C1 value was only found in W23. According to mutation theory any factor in an abnormal state for this organism can induce mutation. Therefore, T. dactyloides genome in the first approximation may be taken as a mutagenic factor. Hence, the possibility to compare values n and C1 is feasible.
The individuals of Zea mays L. are specialized by the main alternative functions: inheritability and variability. The function of Zea genome preservation is realized in inbreds that are low-mutable sensitive and insensitive to mutagen effect. The effect of species preservation is achieved by principally different methods. In sensitive inbreds self-elimination of cells and organisms with distortions in the genome takes place. In insensitive inbreds this effect is realized by active distinction and correction of all distortions or by degradation of an alien genome. Due to the latter, high frequency of haploids in F1 progeny may be expected. The function of genetic variability is concentrated in the lines of moderate insensitivity. These lines are generators of species variability where probably all types of genetic variability are mostly intensified. As for the latter group of inbreds, the majority of maize mutations earlier unknown was induced (MNL 60:70-71, 1986). Genetic analysis showed digenic control of mutability induced by NMU (Genome, 30:159 1988). Probably the systems of unspecific control of the ratio ìvariability:inheritabilityî are conditioned by superloci. The digenic type of control provides preadaptation of the species or evolutionally valuable population to any environment. Let us examine the case of gradual tension increase of mutagen factor in time and space. First of all supersensitive aabb inbreds give a response. Suffering from phenotype depression induced even by small doses of the mutagen they decrease their contribution to the genetic pool of the next generation. It results in an increase of the share of high-mutable aaBB, aaBb genotypes in the population. In case of further increase in tension of the factor the stage of ultimate tension of all types of variability is observed. Search for new approaches to adaptation to changed environment is under way. At last only insensitive low-mutable genotypes AABB, AaBB, AABb, AaBb, improved by new specific factors for stability obtained by crossing with highly variable inbreds, remain. In nuclei of insensitive inbreds adaptively valuable alleles are accumulated and concentrated. The species transits into a genetically closed state where new genotypes mainly occur under the control of the locally specified system. With lessening of mutagen factor tension the species (population) recovers its genetically open evolutionarily valuable preadaptive state (i.e. containing all three types of lines) because of segregation.
Lack of money does not allow the author to thoroughly check all the consequences of the hypothesis. It is necessary for evolutionists, geneticists and molecular biologists to cooperate to realize the project ìGenetic Control of Corn Variabilityî.
Figure.
Sensitivity, mutability and compatibility of maize inbreds. Symbols. Inbreds:
1 - T22, 2 - Gk26, 3 - W23, 4 - A344, 5 - PLS61, 6 - Gb834, 7 - Hy2, 8
- F2. (ÆH/H)/c - unit depression of plant height in M1, where: ÆH
- plant height decrease, H - control plant height, c - NMU concentration,
mM, n - mutation frequency in optimal test variants, C1 - coefficient of
Zea
and Tripsacum dactyloides genome compatibility (see E. P. Erygina
and A. S. Mashnenkov's contribution to this issue).
Return to the MNL 65 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}