University of California
STANFORD, CALIFORNIA
Stanford University
Mu insertion sites at bz-mu1 and bz-mu2: Restriction digests and Southern blotting were used to determine the exact point of insertion of bz-mu2. The element has previously been mapped to within 50 bp (or less) 5' of the XhoII site (AGATCC) in the coding region of Bz-W22. Paradoxically, restriction with Sau3a, an enzyme that cuts within the XhoII site (recognizing GATC) revealed that the 180 bp Sau3a fragment 5' of (and bordering on) the XhoII site does not carry an insertion of any kind. Assuming that the Mu2 element at this site, like the previously sequenced Mu2 element, contains no Sau3a, sites, this apparent contradiction (that the element maps 5' of XhoII but not 5' of Sau3a, ) can be resolved if we assume that the element has inserted within the XhoII site, precisely after the Sau3a site. Because Mu insertion generates a 9 bp repeat, the following sequence should exist at the bz-mu2 allele: ...GCGGAGATC(Mu2)GCGGAGATCC... In this way the insertion of Mu regenerates the XhoII site 3' of the element, while maintaining the Sau3a site 5' (and 3') of the insertion site. We plan to confirm the sequence of the putative 9 bp repeats by cloning and sequencing.
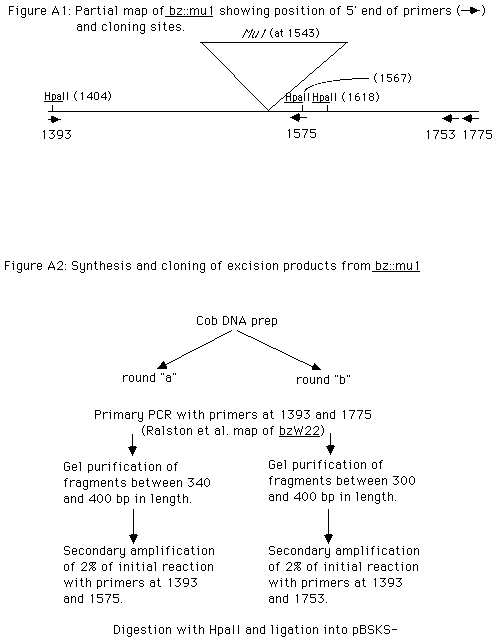
In order to determine the points of insertion of the Mu1 element of the bz-mu1 allele, PCR was used to amplify the bz sequences bordering the insertion. The amplified fragments were then cloned and sequenced. The sequence of the 9 bp repeat, located just 3' of the bronze1 intron, is CAAACAGGG. Remarkably, this is the exact point of insertion of the Mu7 element at the bz-rcy allele reported earlier this year by Schnable, Peterson, and Saedler (MGG 217:459, 1989). There is virtually no possibility that one of these mutable alleles was derived from the other: they carry Mu elements with homology only within the ~~200 bp TIRs, and they were isolated independently (bz-mu1 at Stanford in the summer of 1982, bz-rcy at Iowa State) by laboratories that do not directly exchange materials. The significance of the isolation of two independent insertions at precisely the same site is unclear. The handful of Mutator insertion sites sequenced so far have displayed no homology at the 9 bp repeats. The possibility exists that Mu may have a tendency (but not an absolute requirement) for insertion at sites near to a certain sequence or secondary structure. Only the sequencing of more Mu insertion sites will reveal whether or not the selection of Mu insertion sites is totally random.
Sequence of Mu1 somatic excision sites: In order to determine the nature of the lesion left behind in the host chromosome after Mu excision, we have cloned a number of "empty sites" generated at the bz-mu1 allele. DNA carrying such empty sites can be isolated from germinal revertants, and two excision products of rcy:mu7 from bz generating phenotypically revertant plants (fully colored kernels) have been analyzed by Schnable et al. Unfortunately, this technique for the selection of excision products will yield only those products which generate alleles which cause phenotypic revertants. In order to sequence a wider spectrum of excision products, we used PCR to specifically amplify the products of Mu1 excision from bz present in an immature cob from an active Mutator line homozygous at bz-mu1. If we assume that Mutator activity in cob tissue is roughly the same as that in aleurone (that is, perhaps 5% of all cells have experienced Mu excision from bz--Taylor and Walbot, Genetics 117: 297, 1987--and excision sectors are an average of 20 cells in size), a 2 mg DNA prep (with a haploid genome size of 3 pg) should contain about 20 copies each of 1.7 million independent excision events. Any primary PCR reaction, employing 0.4 µg of chromosomal DNA, should include, on average, 6800 molecules of "empty sites" at bz, most of which are derived from independent excision events.
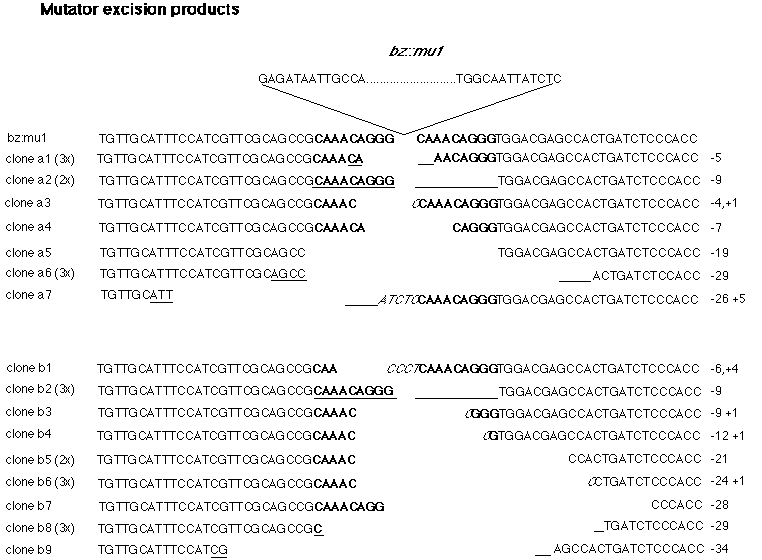
We have found that the Mu element (perhaps due to its TIRs) is itself a very effective block to PCR amplification. Thus, using bz specific primers on both sides of the Mu insertion, specific amplification of excision products was achieved. Because the initial round of synthesis produced DNA in levels insufficient for cloning, a second round of synthesis (with one "nested" primer and one original primer) was undertaken to increase both the quantity and specificity of the products. These were then digested with HpaII as depicted in Figure A, and cloned into pBSKS- for sequencing. The sequence of the excision products is presented below. The number of different excision products cloned and sequenced was not consistent with our estimate of the number of independent excision events per primary PCR reaction. Considering the frequency at which sequences were cloned in duplicate or triplicate (even from independent ligation reactions), either the number of independent excision events present in the original DNA prep was lower (by 3 orders of magnitude!) than that estimated above, and/or the number of independent excision events amplified by the primary PCR reaction was much lower than the initial number of independent products present in the 0.4 µg of chromosomal DNA added to the primary reaction. The presence of an excess of triplicates over duplicates strongly suggests that the different excision products are represented (at the end of the secondary PCR reaction) in different frequencies. Such differential representation in the "pool" of DNAs for cloning might be the result of skewed amplification during the early primary PCR reactions, but it might also represent a tendency by Mutator to delete chromosomal sequences in a nonrandom fashion. In order to distinguish between these two possibilities, the entire protocol (both primary and secondary PCRs) was performed twice on the same cob DNA prep. The second trial produced a different set of clones, with the same excess of triplicates over duplicates. These results suggest that the skewed distribution of excision product types is due to the process of amplification by PCR, rather than to a nonrandom behavior of the Mu element. Clones from trial "a" vs trial "b" are identified as such. Presumably additional repeats of this protocol would continue to yield additional excision products.
Figure
A (continued).
If Mu were to excise precisely,
removing only Mu sequences from the insertion site, two copies of
the 9 bp repeat should be left behind. Of the sixteen different excision
products sequenced, none were of this structure. The excision products
we obtained can be divided into two broad categories. Simple imprecise
deletions of Mu from bz are observed in eleven of the
16 sequenced products. In these products, as in the two sequenced by Schnable
et al., the excision of Mu has resulted in the loss of bz
sequences bordering the insertion site. There is no convincing evidence
for a preference for any particular 5' or 3' endpoint for the deletion.
In fact, in clone a7 the 3' deletion endpoint is 5 to 8 bp within the Mu
element itself. Clone a3 might also be an example of a 3' deletion endpoint
within Mu. On the other hand, a3 might also be the result of the
same repair syntheses which produced the second category of excision products,
the complex imprecise deletions. Clones b1, b3, b4, and b6 all carry
sequences not present in the original bz-mu1 allele, as well as
deletions extending into bz. In three of these four cases the additional
sequence is simply a C residue. In two clones this C residue unambiguously
replaces an A in the original sequence (clones b3 and b6). In a third clone
(b4) the C might replace either an A or a G. The other clone which carries
additional sequences not present in the original bz-mu1 allele is
b1, which carries a CCCT insertion. While it is possible that this insertion,
like the insertions of single C residues, is derived from some sort of
error-prone DNA repair activity, this sequence is closely related to the
3' (AGGG) end of the 9 bp target site repeat. Either the "hairpin loop"
model suggested by Coen, Carpenter, and Martin (Cell 47:285, 1986) or Saedler
and Never's "strand switching" model (EMBO J. 4:585, 1985) for the generation
of post-excision footprints would account for the generation of this 4
bp insertion.
The only clone isolated in both trials is the clone that restores the original sequence of the Bz-W22 allele (a1 and b2). While it is possible that this sequence was amplified as a contaminant in the DNA prep (either from contaminating W22 DNA in the first round of cloning, or contaminating clone a1 in the second), the fact that this excision product appears at approximately the same frequency as other excision products makes this seem unlikely. This (and the other excision products) is not an artifact generated by PCR; extensive attempts at amplification with Mu-containing plasmid templates failed to generate this or any other excision sequences. It is possible that this sequence is produced more often upon excision than any other single excision product. Alternatively, this sequence might be generated (perhaps even in inactive lines) by homologous recombination between the 9 bp repeats, as suggested by Kloeckener-Gruissem and Freeling (MNL, 1989).
The sequences obtained so far of Mu insertion sites show that newly transposed Mu elements carry with them only Mu sequences; the chromosomal sequences from the previous insertion site are not carried along to the new target. In order to generate "clean" elements for transposition, the sequences immediately abutting the element must be nicked precisely at some time during the transposition process. If Mu truly transposes replicatively, in the same fashion as the Mu phage, the first step to transposition should be the generation of single stranded nicks on each side of the element (one to each strand). Assuming that the same enzymes required for replicative transposition are also employed for excision, one would then expect that some excision products would still contain some Mutator sequences, as only one strand of Mu, on each side of the element, is specifically nicked. The structure of clone a7 supports this model, providing evidence that this nicking does not always occur on both strands of both sides of the element.
The amplification and cloning process limited the size and position of the deletions within the excision products (see Figure A). For clone set "a" the maximum clonable deletion size was limited to approximately 35 bp by the gel purification of the primary PCR reaction. The range of fragment sizes gel purified for clone set "b" was broader, and the nested primer for secondary synthesis set farther 3' of the insertion site, permitting the cloning of excision products carrying deletions of up to 75 bp. Even within these limits, the size and position of the deletion present in each of the excision products does not appear to be totally random. Smaller deletions occur more frequently than large ones; no deletion extended more than 26-29 bp 5' or 26 bp 3' of the insertion site. If the position of the deletion end points is truly random (i.e., entirely unrelated to the position of the Mu insertion) such a distribution might be expected for clones from set "a", but certainly a broader range of deletion sizes would be expected for clone set "b". The fact that deletion endpoints cluster near the point of Mu insertion supports the model suggested above�that the initial nicks are generated specifically at the point of insertion of Mu. The deletions might then be extended by exonucleolytic degradation.
This semirandom nature of the distribution
of deletion endpoints in the excision products seems at first to suggest
that the excision process of the Mu element includes no mechanism
for the restoration of the integrity of the host chromosome�-that the element
simply hops out and leaves the two broken ends behind. If this were so,
however, one would expect every chromosome which has experienced an excision
event to be equivalent to a broken chromosome, such as those generated
by gamma irradiation. This does not seem to be the case. Recent data presented
by Rowland, Robertson, and Strommer (Genetics 122:205, 1989) have demonstrated
that Mu activity, or at least activity in the female germline, does
not frequently induce chromosome breakage. Our own unpublished data support
these results. The sequenced footprints of other transposable elements
of maize (with the exception of certain "chromosome breaking" elements)
suggest that these elements behave similarly to Mu; excising imprecisely,
and yet not causing chromosome breakage. Thus any model for transposable
element excision/transposition must explain this ability to rejoin, however
imprecisely, the 5' and 3' ends of the empty site.
Return to the MNL 64 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}