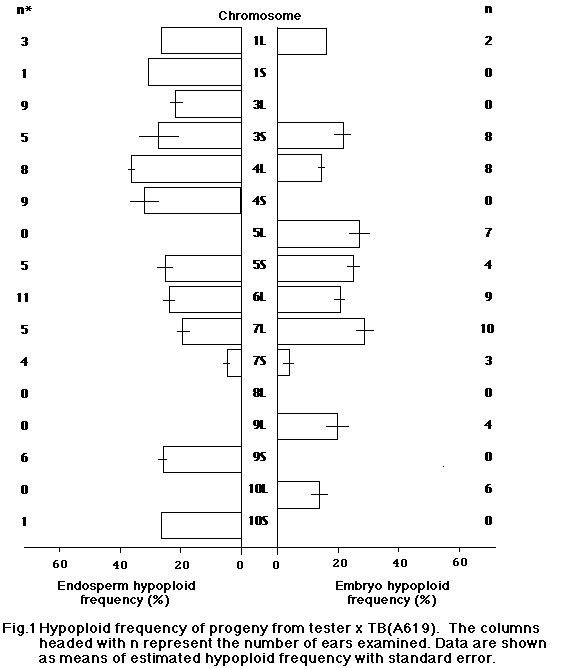
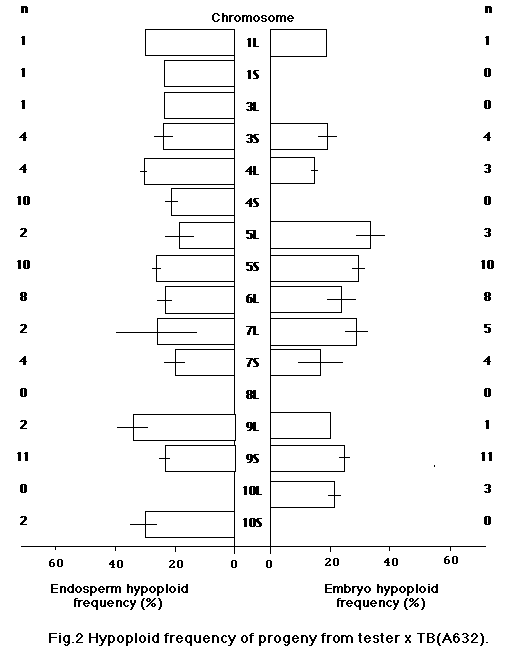
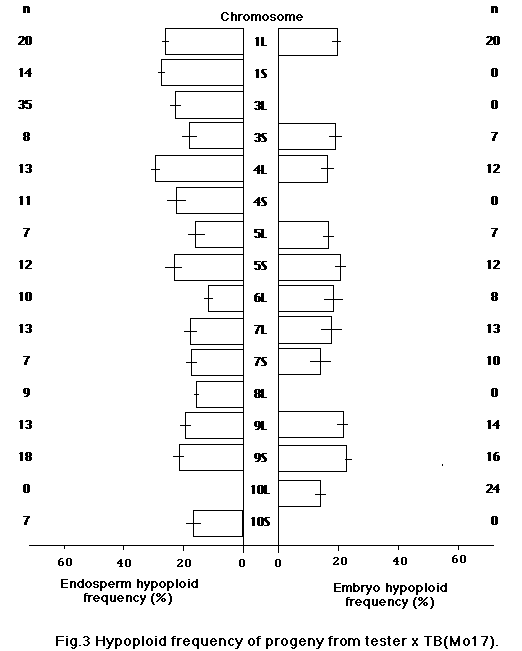
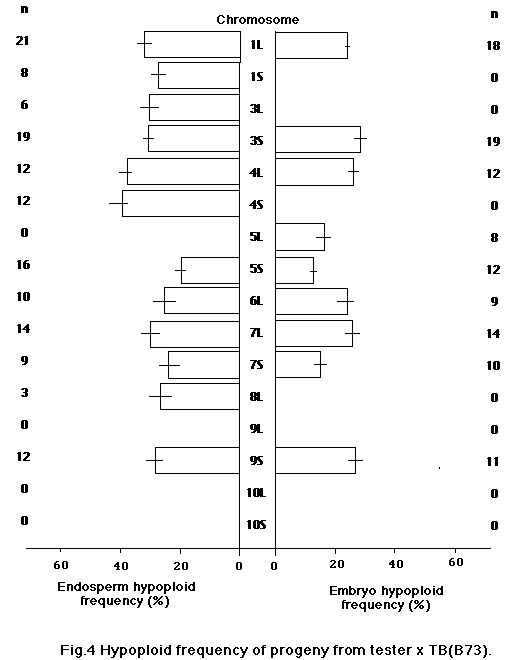
In this study we attempted to evaluate the effect of genetic background of B-A translocations on the frequency of hypoploid progeny. We used 17 B-A translocations which were backcrossed up to five times with four inbred lines, A619, A632, Mo17 and B73. They were crossed on appropriate testers in 1989, and the hypoploid frequency of endosperm and embryo was determined from the recessive characters of kernels or of seedlings on sandbench tests. Materials are shown in Table 1.
Table 1. List of B-A translocations
and testers.
|
|
|
|||||
| B-A translocation |
|
|
|
|
|
|
| TB-1La |
|
|
|
|
|
|
| TB-1Sb |
|
|
|
|
|
|
| TB-3La |
|
|
|
|
|
|
| TB-3Sb |
|
|
|
|
|
|
| TB-4Lc |
|
|
|
|
|
|
| TB-4Sa |
|
|
|
|
|
|
| TB-5La |
|
|
|
|
|
|
| TB-5Sc |
|
|
|
|
|
|
| TB-6Lc |
|
|
|
|
|
|
| TB-7Lb |
|
|
|
|
|
|
| TB-7Sc |
|
|
|
|
|
|
| TB-8Lc |
|
|
|
|
|
|
| TB-9Lc |
|
|
|
|
|
|
| TB-9Sd |
|
|
|
|
|
|
| TB-10L19 |
|
|
|
|
|
|
| TB-10Sc |
|
|
|
|
|
|
For each ear 50 kernels were used to estimate the hypoploid frequency in progeny. Hypoploid frequency was calculated from the number of kernels or seedlings that showed recessive characters. As shown in Figures 1-4, hypoploid frequency varied depending on the background and the B-A translocation. We found a significantly lower hypoploid frequency for some B-A translocations with Mo17 background. It appears that the background of B-A translocations affects the transmission rate. Also, hy-poploid frequency varied for each TB significantly. However, the cause of the difference between TB's is not clear, since testers were different for each TB.
Table 2. List of testers.
|
|
Genotype and background |
|
|
bz2 R-r b pl [W22b1x W23] |
|
|
bz2 R-r b pl [W23 x W22b] |
|
|
[B73 x A632] x (d1 + / d1 cl1) |
|
|
[Mo17 x A619] x (d1 + / d1 cl1) |
|
|
[A632xMo17] x (d1 + / d1 cl1) |
|
|
[B73 x A632] x (d1 + / d1 cl1) |
|
|
c2 R-r b pl [K55 x W23} |
|
|
c2 R-r b pl [W22b x K55] |
|
|
c2 R-r b pl [W23 x W22b] |
|
|
a2 R-r b pl [W22b x K55] |
|
|
a2 R-r b pl [W23 x K55] |
|
|
a2 R-r b pl [K55 x W23] |
|
|
a2 R-r b pl [W22b x W23] |
|
|
y1 l12 / y1 + |
|
|
[B73 x A619] x o5 gl1 |
|
|
[Mo17] x o5 gl1 |
|
|
([B73xMo17] x (+/vp9)) selfed |
|
|
[B73 x Mo17] x o2 v5 |
|
|
sh bz wx R-r/R-scm [W23 x W22b] |
|
|
sh bz wx R-r/R-scm [W22b x W23] |
|
|
bz R-r/R-scm [W23 x W22b] |
Table 3. Endosperm and embryo classification
of progeny from tester X TB
|
|
|
||||
| TB background |
|
|
|
|
|
| A619 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| A632 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| Mo17 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| B73 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
2Recessive frequency between kernels
and seedlings was examined by t-test. T-value for difference of recessive
percent was significantly different at 0.05 level(*) or 0.01 level (**).
Since the genotype of the tester for TB-6Lc consists of a homozygous endosperm
marker and heterozygous seedling marker, percentages of recessive seedlings
were doubled and examined by the t-value.
TB-7Sc with A619 background showed extremely low hypoploid frequency. However, this TB was backcrossed only once with inbred A619 and the others, so that it is difficult to determine the effect of inbred background.
For several of the TB's, the hypoploid frequency was determined for both kernel and seedling characters. Carlson (1969) reported that the occurrence of preferential fertilization is dependent on testers. To analyze the tendency for preferential fertilization, hypoploid frequency was examined for each tester. Information on testers is shown in Table 2, and data are given in Table 3.
With TB-4Lc, preferential fertilization occurred without regard to inbred background. Although Carlson (1969) found that some testers generate equal fertilization, all of our testers showed preferential fertilization for this chromosome. However, the difference in recessive frequency between endosperm and embryo seemed to be altered depending on the tester. This might be more clear after additional investigations.
Preferential fertilization also occurred
with TB-1La in Mo17 and B73 background, except for 1L1 tester when it was
pollinated by B73 background. This indicates the possibility that tester
and TB background generate the preferential fertilization together.
Return to the MNL 64 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}