The approach might be called 'tertiary monosomy', as it employs r-x1-induced nondisjunction to generate losses of a translocated chromosome. The design is to cross the translocation stock onto r-x1 stock, and to cross the F1 plants by recessive seedling or plant markers that are located on one of the translocated chromosomes. Selected recessive plants will include ones that have lost the translocated chromosome on which the test gene is located, and the translocated segment of the other chromosome. Diagrammatically, the following represents the F1 for a translocation between chromosomes 1 and 2, in which the gene A, affecting a plant trait, is near the end of one arm of chromosome 1:
1111o1111111111111A11 222222o22222222222 r/r-x1
1111o1111112222222 222222o22221111111A11
This F1 will generate eggs deficient for A for either of two reasons. Either the whole chromosome 1 is lost, or a translocated chromosome bearing that segment of 1 translocated with 2. The progeny from crosses by a will express the recessive gene, and will be either the monosomic or the tertiary monosomic. When RFLP probing is done for loci on chromosomes 1 and 2, the deficiencies in each of the two chromosomes can be defined, not only parsing both chromosomes for the deficiency they generate but pinpointing the translocation breakpoint on each of the two chromosomes.
We are now screening a pilot study, using a series of 13 wx translocations crossed onto r-x1 and testcrossed with yg2 and Bf1 (and several other markers on chromosome 9). Tertiary monosomics occur, albeit at a low rate (1 per 1000). Leaf samples are to be collected from them when they grow large enough, and RFLP probing will be carried out. An earlier pilot produced a small sample of cases, from which preliminary probings showed some of the expected events.
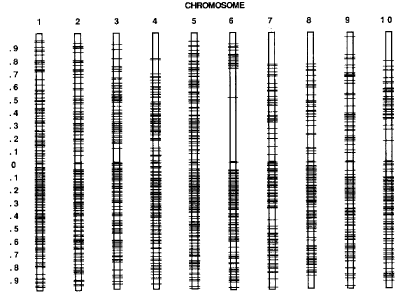
Figure. Breakpoints of the reciprocal translocations.
In the summer of 1989, we planted samples
of all 879 of the translocations from the Stock Center (obtained with the
enthusiastic help of Earl Patterson) and achieved crosses of over 800 onto
r-x1
stocks. As testcrossing and screening proceed, we hope to be able to define
as many as possible of the deficiencies produced with these translocations.
The accompanying figure shows the distribution of cytologically determined
breakpoints across the genome. Whether 800 translocations (1600 breakpoints)
can be defined, or only as few as 400 or 500, each translocation characterized
will represent reproducible physical breakpoint locations on two chromosomes,
at a wide distribution of physical locations.
Return to the MNL 64 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}