BLACKSBURG, VIRGINIA
Virginia Polytech. Institute & State
Univ.
Proteolytic activity in coleoptile extracts
causes artifactual ß-glucosidase multiplicity
--Asim Esen and Cumhur Cokmus
Maize ß-glucosidases (ß-D-glucoside
glucohydrolase, EC 3.2.1.21) isolated from inbred lines display multiple
electrophoretic variants. These variants appear in a time-dependent manner
during storage, most notably at acidic pHs (below pH 6.0) and in the presence
of a reducing agent. The enzyme was extracted from the coleoptiles of 6-day-old
seedlings with a variety of aqueous buffers and assayed for activity. Zymograms
were obtained by incubating the gels with the synthetic substrate 6-bromo-2-naphthyl
ß-D-glucopyranoside after electrophoresis. The pHs of the crude enzyme
preparations (extracts) were adjusted to vary from 3.0 to 10.0, and the
preparations were incubated at different temperatures (-30, 4, 25, and
37
C) for varying lengths of time in the presence and absence of the reducing
agent 2-mercaptoethanol (2-ME) and assayed for activity and changes in
zymogram patterns. The results showed that the enzyme lost activity at
higher temperatures (25 and 37 C) and at pHs below 4 and above 9. Similarly,
the number of charge variants resolved by electrophoresis increased during
incubation, especially at higher temperatures. These novel variants had
increased anodal mobilities, and could be resolved into 3 to 5 distinct
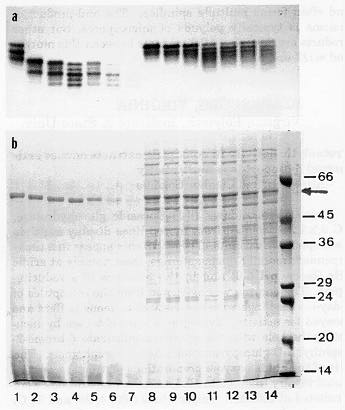
bands after incubation at pHs between 4 and 6 (Fig. 1a, lanes 1-6). When
polypeptide profiles of the extracts were analyzed by sodium dodecyl sulfate
polyacrylamide gel electrophoresis (SDS-PAGE) after the above-mentioned
treatments, it was observed that the staining intensity of individual polypeptides
decreased and there was a complete loss of some polypeptides at pHs between
4 and 6 in a time-dependent manner under reducing conditions, especially
after incubation at 25 and 37 C (Fig. 1b, lanes 1-7). An example of the
results is shown in Fig. 1. Figure 1a shows the changes in ß-glucosidase
zymograms of the coleoptile extract from inbred K55 made with 25 mM sodium
acetate buffer (pH 5.0) containing 35 mM 2-ME. The extract (final pH 5.2)
was incubated at two different temperatures (25 and 37 C) for 6, 12, and
24 hours, respectively. It is evident that zones (bands) of ß-glucosidase
ac-tivity increase in number, and novel anodal bands appear as cathodal
bands disappear or decrease in intensity in a time- and temperature-dependent
manner (Fig. 1a, lanes 2-6). When the pH of the same extract was adjusted
to 7.0 before treatments, the shifts from cathodic to anodic zones of activity
slowed substantially and the anodic shift produced mostly a diffuse zone
of activity instead of distinct bands (Fig. 1a, lanes 9-14). At both pHs,
the highest enzyme activity and the least anodal shift in zymograms was
in the samples stored at 4 C for 24 hours (Fig. 1a, lanes 1 and 8). Figure
1b shows the changes in polypeptide profiles of the same samples used to
develop the zymograms in Figure 1a. It is apparent that the cause of the
changes in zymograms at pH 5.2 is proteolysis because the size and intensity
of the ß-glucosidase polypeptide (60 kD band, arrow) decreases in
a time- and temperature-dependent manner (Fig. 1b, lanes 2-7). Moreover,
essentially complete proteolysis of polypeptides other than the ß-glucosidase
monomer is evident from (1) the absence of any bands in lanes 1-7 (Fig.
1b), and (2) the presence of a diffuse, low molecular weight Coomassie
blue stained zone at the anodic end (bottom) of the gel. In contrast, little
or no proteolysis is evident in samples whose pH was adjusted to 7.0 (Fig.
1b, lanes 8-11), except some slight proteolysis at 37 C (Fig. 1b, lanes
12-14). We were able to inhibit the proteinase active at acidic pHs with
antipain, leupeptin and the alkylating agent iodoacetic acid, but not the
one active at neutral or alkaline pHs. Based on these data, it was concluded
that maize ß-glucosidase multiplicity observed in zymograms of inbred
lines is an artifact of proteolysis. An SH-proteinase active at acidic
pHs and requiring a reducing agent (e.g., 2-ME) for activation was primarily
responsible for artifactual ß-glucosidase multiplicity. Another class
(undefined) of proteolytic activity was found to be responsible for producing
charge multiplicity above pH 6; but this multiplicity appeared mostly as
a broad and diffuse zone instead of distinct bands. Using buffers of neutral
or alkaline pH or excluding reducing agents from the extraction medium
and performing all the operations at 0-4 C appear to be effective strategies
against the SH-proteinase.
Figure
1. Time-course of the changes in ß-glucosidase zymograms (a)
and polypeptide patterns (b) after extraction with 25 mM Na-Acetate, pH
5 / 35 mM 2-ME. The extract (pH 5.2) was incubated at 25 and 37 C for 6,
12, and 24 hrs. Lane 1, 4 C control (24 hrs); lanes 2-4, after 6, 12, and
24 hrs, respectively, at 25 C; lanes 5-7, after 6, 12, and 24 hrs, respectively,
at 37 C. Lanes 8-14, after the pH of the extract was adjusted from 5.2
to 7. Lane 8, 4 C control (24 hrs); lanes 9-11, after 6, 12, and 24 hrs,
respectively, at 25 C; lanes 12-14, after 6, 12, and 24 hrs, respectively,
at 37 C. The last lane (unnumbered) in b includes marker proteins ranging
in size from 14 to 66 kD. The arrow on right points to the 60 kD polypeptide
(ß-glucosidase monomer).
Please Note: Notes submitted to the Maize Genetics Cooperation
Newsletter may be cited only with consent of the authors
Return to the MNL 64 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}