Louisiana State University
ST. PAUL, MINNESOTA
University of Minnesota
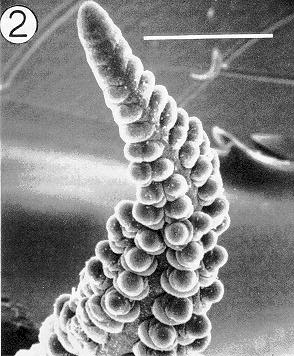
An examination of more than 50 developing tassels and ears of Argentine pop revealed that, in the majority of cases, early primordia are produced in two ranks by the apical meristem of the inflorescence. Figure 1 illustrates a young tassel primordium oriented such that one rank of initials (lateral primordia) is in face view on the distal end of the axis. These initials enlarge laterally, then bifurcate to form paired primordia. Such bifurcation is clearly visible near the middle of the axis in this figure. However, rather than initiating a pair of spikelets (one sessile, one pedicellate) as would be expected in teosinte inflorescences or the lateral tassel branches of maize, these paired primordia bifurcate a second time. The result of this second bifurcation is seen clearly in Figure 2, which illustrates an ear primordium at a slightly later stage of development. Above the middle of this inflorescence, primordia are in two ranks; however, below the midpoint, there are four ranks (8 rows) of spikelet primordia as a result of the second bifurcation. Two of these ranks are visible and two additional ranks are out of view behind the axis. Thus, as a result of the second bifurcation, an inflorescence meristem that initially produced only two ranks of primordia (i. e., distichous) becomes one with four ranks of spikelet primordia (i. e., polystichous).
Figure 1. A young tassel primordium of Argentine popcorn. The specimen is oriented such that one rank of initials on the distal end of the axis is in face view. These initials are distichously arranged as would be found in teosinte or lateral branches of the maize tassel. Bar = 50 µm.
Figure 2. An ear primordium of Argentine popcorn. The specimen is oriented such that the plane of the initial bifurcation is in face view. The older (proximal) initials have undergone a second bifurcation resulting in four ranks of spikelet pairs. Two of these ranks are hidden behind the axis. Bar = 500µm.
It is also apparent in Figure 2 (although perhaps most clearly to the trained eye) that the ranks of spikelet primordia are not exactly opposite each other along the upper portion of the axis, but are shifted toward the abaxial side of the developing ear. Even at the stage when the inflorescences first become eight rowed, they are bilaterally symmetrical with the rows shifted toward the abaxial side of the axis (not shown). However, subsequent enlargement of the spikelets displaces the rows into a radially symmetric arrangement as illustrated by the polar view of a slightly older central tassel spike (Fig. 3). These observations are consistent with the derivation of the maize ear and central tassel spike from distichous, bilaterally symmetrical inflorescences such as those of teosinte.
From these observations, we have formulated a model for the origin of polystichy in maize. This model takes into account the fact that virtually all of the Andropogonoid grasses (the group to which maize belongs) have distichously arranged sessile-pedicellate pairs of spikelets. As such, the inflorescences of this enormous group of grasses must share a common developmental program by which the apical meristem first produces distichously arranged primordia, each of which bifurcates to produce paired (sessile-pedicellate) spikelet primordia. In this framework, the origin of polystichy in maize can be seen as a change in the timing of the developmental program controlling bifurcation of the initials. If the execution of this program is simply modified (prolonged) to allow one additional cycle of bifurcation, a four-ranked, eight-rowed inflorescence is produced, this the most primitive condition in cultivated maize. This is apparently what happens in Argentine pop. Thus, "twisting" as suggested by Collins would not be involved in the origin of polystichy, which is consistent with the fact that we see no evidence for twisting in the development of Argentine pop inflorescences. Other modifications of the program controlling bifurcation would be required to produce 10-, 12- or 14-rowed inflorescences. Ontogenetic analyses of maize varieties with 10- or higher-rowed inflorescences should provide insights into the nature of these modifications. Additional studies should also clarify the developmental basis of the production of inflorescence branches, which are also the products of bifurcation of the developing inflorescence primordia and thus also occur in pairs. We are currently exploring these issues and testing the validity of our model for the origin of polystichy.
Figure
3. A polar view of a developing central tassel spike of Argentine popcorn.
This specimen shows how enlargement of the spikelets displaces the rows
into a radially symmetrical arrangement. Bar = 500 µm.
Return to the MNL 64 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}