Cytoplasmic reversion in M825 and 38-11 is characterized by the loss of the S1 and S2 molecules and rearrangements of S2 or S1 homologous sequences in the main mitochondrial genome, respectively. The S1 and S2 targets for recombination, specifically the IR-homologous sequences present in the sigma-R and psi-R regions of the main mitochondrial genome, are not affected (Theor. Appl. Genet. 76:609, 1988). However, linear chromosome termini which have homology to the R repeat, the 2kb sequence common to regions of the main mitochondrial genome termed sigma' and psi' and that contains the IR-homologous sequence, are lost (Theor. Appl. Genet. 76:609, 1988; Phil. Trans. R. Soc. Lond. Ser. B 319:149, 1988; Nature 310:292, 1984).
In contrast, cytoplasmic reversion in the inbred line WF9 is characterized by the maintenance of S1 and S2 in equimolar amounts and the lack of rearrangements involving S1 and S2 homologous sequences in the main mitochondrial genome (Theor. Appl. Genet. 75:659, 1988). Interestingly, it is the S1 and S2 targets for recombination that are lost (Theor. Appl. Genet. 76:609, 1988).
To further characterize the molecular aspects of cytoplasmic reversion involving the R repeat, two sibling, WF9-RD cytoplasmic revertants (cyto-revs) were converted to the M825 nuclear background by backcrossing with M825 as the recurrent male parent. After the first backcross to M825, some exceptional sterile plants were obtained from one of these cyto-rev sources. Both the fertile and sterile plants were propagated by continued backcrossing to M825.
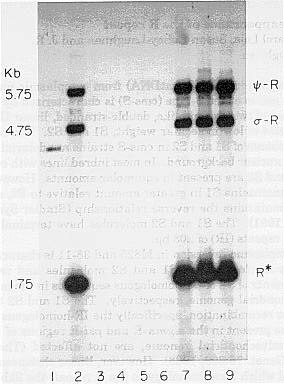
MtDNA from WF9-N x M82510, WF9-RD (sterile), WF9-RD cyto-rev 81-47-13 (fertile), WF9-RD cyto-rev 81-47-15 (fertile), 81-47-13 x M8256 (fertile), 81-47-15 x M8256 (fertile), 81-47-15 x M8256 (sterile), 81-47-15 x M8257 (sterile), and 81-47-15 x M8256 (sterile; independent case) was digested with BamHI and electrophoresed on a 1% agarose gel and blotted onto a nylon membrane. The Southern blot was probed with IS1E5, a clone specific for the R repeat. Consistent with the findings of Small et al. (Theor. Appl. Genet. 76:609-618, 1988), the sibling WF9-RD cyto-revs lost the S1 and S2 target sites for recombination (See Figure 1, lanes 3 and 4.). However, in contrast to the WF9-S cyto-rev described by Small et al., the chromosome termini (R*) were also lost.
Figure 1. Hybridization of IS1E5 (specific for the R repeat) to a Southern blot of BamHI-digested mtDNA. Lanes: (1) WF9-N x M82510 (2) WF9-RD (sterile) (3) WF9-RD cyto-rev 81-47-13 (fertile) (4) WF9-RD cyto-rev 81-47-15 (fertile) (5) 81-47-13 x M8256 (fertile) (6) 81-47-15 x M8256 (fertile) (7) 81-47-15 x M8256 (sterile; independent case from that in lanes 8 and 9) (8) 81-47-15 x M8257 (sterile) (9)81-47-15 x M8256(sterile).
Even after conversion of the sibling WF9-RD cyto-revs to the M825 nuclear background, the male-fertile plants neither lost the S1 and S2 episomes nor regained the target sites for recombination normally retained by M825 cyto-revs (lanes 5 and 6). Reversion to fertility in WF9, however, did not affect the ability of S1 and S2 to change their relative copy number in response to the M825 nuclear background. The relative abundance of S1 and S2 was that characteristic of M825 (MNL 62:109, 1988).
The exceptional sterile plants obtained
after the first backcross of the sibling WF9-RD cyto-revs to M825 that
were continually backcrossed also had S1 and S2 in the relative abundance
characteristic of M825. However, these sterile plants apparently regained
the S1 and S2 target sites for recombination and the R* linear chromosome
termini (lanes 7-9). This observation is similar to the findings of Wang
and Gengenbach (Maydica 34:217, 1989), who noted the appearance of the
S-atpA-3
gene from progenitors in which it was not detected. A plausible explanation
for the loss and reappearance of the R repeat is the reduction of the sequence
to undetectable levels followed by selective amplification.
Return to the MNL 64 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}