Because we obtained low yields of amyloplasts and the amyloplast isolation procedure requires that amyloplasts contain starch grains (13 to 14 dap and older), we decided to use plastid membranes isolated from the microsomal fraction for the production of polyspecific polyclonal antibodies. Membranes for antibody production were isolated from 50 to 100 g of A188 and A619 x W64A endosperm 15 to 17 dap. Two chickens were injected with envelope membrane vesicles of A188 (800 ug of protein total/chicken) and two with vesicles of A619 x W64A amyloplast membranes (1 mg of protein total/chicken). Membrane vesicles were emulsified with complete Freunds adjuvant for the first injection and with incomplete Freunds for three boost injections. Antibodies were isolated from eggs after the fourth injection.
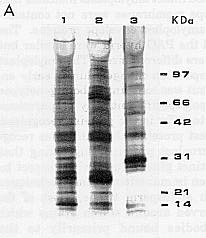
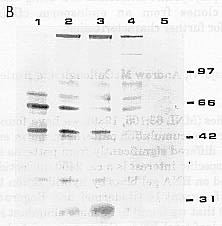
Figure 1. (A) Plastid envelope membrane proteins separated by 10-15% gradient SDS-PAGE. Envelope proteins from A188 12 dap endosperm microsomal fraction (Lane 1), A188 15 dap isolated amyloplasts (Lane 2), and leaf chloroplasts (Lane 3). Lane 1 has 67% of the total protein run in Lanes 2 and 3. (B) Endosperm plastid envelope membrane polyspecific antibodies reacted with envelope proteins separated by 10% SDS-PAGE. Envelope proteins from microsomal fraction of A188 8 dap (Lane 1) and 12 dap (Lane 2), and A619 X W64A 16 dap (Lane 4), from isolated amyloplasts of A188 15 dap (Lane 3), and from A188 leaf chloroplasts (Lane 5). An equal amount of protein was run in each lane.
Endosperms were collected and membranes isolated from 12 to 25 g of A188 (white endosperm) 8, 9, 10, 11, 12 and 16 dap and from an early season PAG hybrid (yellow endosperm) 8, 10, 12, 14 and 16 dap. For comparison, chloroplast envelope membrane proteins were isolated from 7 to 10-day-old maize seedlings using the method of Keegstra and Yousif (Meth. Enzymol. 118:316, 1986). Intact chloroplasts were isolated, purified on a Percoll gradient, lysed osmotically and the thylakoid membranes removed with centrifugation and the envelope vesicles purified on a sucrose step gradient. The protein fractions were analyzed on 10 to 15% and 8 to 25% acrylamide gradient SDS-PAGE.
Compared to chloroplast envelope membrane
proteins, amyloplast envelope membrane protein profiles were quite different
with only a few major bands with similar mobility detected (Fig. 1). The
similarity of the patterns for envelope membranes isolated from the microsomal
fraction and from isolated intact amyloplasts indicates that the amyloplast
envelope membranes were not contaminated with many non-amyloplast envelope
proteins. The patterns for A188 and the PAG hybrid were similar but some
major proteins were different sizes. The amyloplast membrane protein composition
changed during early endosperm development but was constant in both genotypes
after day 12. Three polyspecific antibody preparations were tested and
found to differ in the proteins recognized. Compared to amyloplast proteins,
the antibodies recognized only a few chloroplast proteins, suggesting that
amyloplasts are a distinct plastid form with respect to envelope proteins.
When immunofluorescence techniques were used with fresh endosperm tissue,
most of the fluorescence was observed around starch grains which indicated
that antibodies bound primarily to the amyloplast envelope membrane, but
some weak fluorescence also was observed from the plasma membrane. These
polyspecific antibodies are being used to isolate cDNA clones from an endosperm
cDNA expression library for further characterization.
Return to the MNL 64 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}