Carleton College
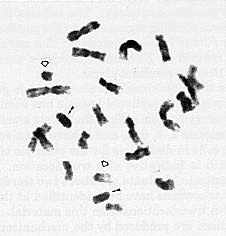
When root-tip chromosomes were examined to ascertain B chromosome number in test cross progeny, many plants were found with two telocentric fragments in addition to the B chromosomes which were present in most plants. One of these additional telocentric fragments is twice the length of the other and is hard to discern from a B chromosome in some mitotic spreads. However, in many cells the B chromosomes are more heterochromatic than either of the newly produced telocentric fragments, indicating that the new telocentrics are derived from an A chromosome (Figure 1).
Figure 1. Mitotic chromosomes from plant containing two B chromosomes (small arrows) and both telocentric fragments (large arrows). The large telocentric fragment, though similar in length to a B chromosome, is clearly less heterochromatic and displays a separation of sister chromatids not seen in B chromosomes.
Karyotype analysis of plants with telocentric fragments indicated that the centromere breakage probably occurred in chromosome 3. Crosses to translocation stocks and marker gene stocks are being carried out to confirm this identification.
Pollen mother cells were examined from plants containing the two telocentric fragments and their normal homolog. Our observations indicate that each telocentric fragment pairs with one arm of the normal homolog and that subsequent prophase I centromere orientation generally yields one of two metaphase I trivalent configurations. A Type I configuration is produced when the centromeres of the two telocentric fragments align toward one pole and that of the normal homolog aligns toward the other pole. A V-shaped trivalent with three centromere spindle attachments is formed. This trivalent is consistently observed off the metaphase I plate (Figure 2). The two centromere spindle attachments of the telocentric fragments appear to provide for a stronger poleward force than does the single centromere spindle attachment of the normal homolog.
Figure 2. Type I trivalent configuration in metaphase I of meiosis in a plant containing both telocentric fragments and no B chromosomes. Both fragments are migrating to the same pole with opposite orientation of the normal homolog. The displacement of this trivalent from the metaphase I plate is common.
Figure 3. Type II trivalent configuration in metaphase I of meiosis from the same plant as in Figure 2. The two telocentric fragments are proceeding to opposite poles with the normal homolog stretched between them. Spindle attachment of the normal homolog is not apparent.
A Type II configuration is formed when the centromeres of the telocentric fragments align toward opposite poles. This alignment appears to preclude attachment of the normal homolog's centromere to the spindle and produces a rod-shaped trivalent in which the normal homolog is stretched between the telocentric fragments (Figure 3).
This breakage event could provide excellent
material for the study of telocentric chromosomes and centromere structure
and function as well as centromere orientation during meiosis. We recently
created a stock that is homozygous for the broken chromosome with 2n =
22. It is still unclear what relationship B chromosomes may have had to
the chromosome breakage event. Additionally, it is unclear how colorless,
sectored, and mottled aleurones of progeny in test crosses are related
to the telocentric fragments. Although it is easy to understand how these
may be produced by instability of the large telocentric fragment in crosses
to a1 testers if the telocentric fragments are actually the arms
of chromosome 3, it is more difficult to provide an explanation for these
exceptional kernels in crosses to c1 testers.
Return to the MNL 64 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}