Illinois State University
B chromosomes are accessory chromosomes of unknown origin which are present in certain maize lines. They have been shown to alter intergenic recombination, intragenic recombination, nondisjunction, and to interact with chromosome knobs (See Carlson, Annu. Rev. Genet. 16:5-23, 1978).
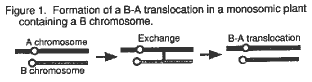
Because exchange has been found to take place between segments of nonhomologous A chromosomes which lack homologs in maize haploids (Weber and Alexander, 1973), we felt that it was possible that exchange might also occur between A chromosomes and B chromosomes in plants where both of these chromosome types were present as univalents. To determine if this could occur, we generated maize plants which were monosomic and contained one B chromosome; and we examined the progeny of these plants for the expected products of recombination between these nonhomologous chromosome types.
Inbred W22 plants with the r-x1 deficiency were crossed by the inbred, Black Mexican, which contained B chromosomes. From this, we derived plants which contained one univalent A chromosome (a monosomic) and one univalent B chromosome as described below. If the nonhomologous univalent A chromosome paired with the nonhomologous univalent B chromosome in these plants and recombination occurred between the two nonhomologous univalent chromosomes as shown in Figure 1, a B-A translocation would be produced. If nondisjunction of the B-A chromosome then took place at the second microspore division, a mature pollen grain with one hypoploid sperm (which does not contain the B-A chromosome) and one hyperploid sperm (which contains two copies of the B-A chromosome) would be produced. If a pollen grain of this type fertilized a plant which was homozygous for a recessive endosperm marker which is located distal (between the breakpoint and the telomere on that chromosome arm) to the breakpoint on the A chromosome and the hypoploid sperm fertilized the polar nuclei, a kernel would be produced which would express the recessive phenotype of this marker in its endosperm. This kernel would also be hyperploid for this B-A translocation in its embryo (that is, it would contain the A-B chromosome and two copies of the B-A chromosome; therefore, it would contain the complete B-A translocation). Thus, several different events must each occur to produce B-A translocations by this mechanism.
To determine if these events could occur, we generated R/r-x1 plants which contained two B chromosomes. These plants were then crossed as female parents by Mangelsdorf's multiple chromosome tester (which is bm2; lg, a, su, pr, y, gl, j, wx, and g on chromosomes 1-10 respectively). Progeny of this cross expressing the gl, j, or g mutant phenotypes were identified as presumptive monosomics for chromosomes 7, 8, and 10 respectively, and plants expressing the distinctive phenotypes characteristic of plants monosomic for chromosomes 4 and 6 were also identified as presumptive monosomic-4 and -6 plants respectively. Nearly all of the progeny of this cross (including the presumptive monosomics) would also contain one B chromosome. Plants larger than typical of these monosomic types were discarded (which often contain deficiencies); thus, most of the presumptive monosomics would have been monosomics.
In order to determine if B-A translocations are produced by the mechanism indicated above, pollen from these monosomic plants containing one B chromosome was placed on silks of plants which contained a recessive marker mutant which is expressed in the endosperm as follows. Monosomic-4 plants were crossed to c2 females, monosomic-6 to su2 females, monosomic-7 to o2 females, monosomic-8 to pro females, and monosomic-10 to y9 and o7 females. Ears produced from these crosses were screened for exceptional kernels which expressed the recessive mutant marker provided by the female tester parent. Each kernel which expressed the marker mutant phenotype was a potential isolate of a new B-A translocation with a breakpoint between the centromere and the mutant marker.
These exceptional kernels were then planted in a field nursery, and sporocyte samples were taken from the resultant plants for cytological analysis. Each of these plants was selfed and testcrossed onto plants which contained the original kernel marker to identify contaminants (which would be homozygous for the mutant marker).
They were also crossed as male parents onto plants carrying recessive seedling markers that mapped near or distal to the original kernel marker as follows: exceptional c2 kernels produced by monosomic-4 plants were crossed to gl3, gl4, and dp females; exceptional su2 kernels produced by monosomic-6 plants were crossed to py females; exceptional o2 kernels produced by monosomic-7 plants were crossed to v5 and vp9 females; exceptional pro kernels produced by monosomic-8 plants were crossed to v21 and gl18 females, exceptional y9 kernels produced by monosomic-10 plants were crossed to oy, and exceptional o7 kernels produced by monosomic-10 plants were crossed to l, and w2 females. Progeny of these crosses were planted in a sandbench and scored for the presence of the seedling markers. Expression of the seedling marker was taken as a positive indicator for the presence of a B-A translocation with a breakpoint in the A chromosome which is proximal (between the gene and the centromere) to the endosperm marker mutant.
Because the pachytene cytology was poor in these plants, the plants produced by the exceptional kernels were also crossed by KYS, an inbred with exceptionally favorable pachytene cytology. The isolates that showed unambiguous seedling segregation in the sandbench plantings were pursued more vigorously. The self of each isolate was crossed by KYS and sporocytes were collected from 5 to 15 of the resultant plants.
At this time, diakinesis in plants which germinated from the original exceptional kernels has been analyzed. Also all sporocytes from selfs of the isolate crossed by KYS from monosomic 2, 4, 6, 7, and 8 have been screened by observing cells in diakinesis for the presence of a ring or chain which is indicative of a translocation. Materials from monosomic 10 plants are about two thirds complete. Seven translocations have been identified at this time including B-A translocations from this material. Thus, B-A translocations are produced by the mechanism described above. It will be extremely interesting to determine if the breakpoints are randomly distributed or at specific positions on the chromosomes.
Much to our surprise, A-A translocations
have also been identified in this material. Some of these A-A translocations
possessed a univalent B chromosome and others did not. Weber (unpublished)
has also recovered several A-A translocations in progeny of plants containing
the r-x1 deficiency. From these results, we can add the production
of translocations to the list of effects known to be caused by the r-x1
deficiency.
Return to the MNL 64 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}