Five tassels of each genotype: 1) homozygous recessive ms-si and 2) heterozygous ms-si (WT), were harvested at random every third day from plants in the field nursery, beginning at the stage of spikelet primordia until anthesis (displayed by WT only). From the mid-portion (one third) of the main rachis, five spikelets (random mixture of sessile and pedicellate) were removed for measurements including length of outer glumes; lodicules and stamens (excluding any silk-like structures, SLS) of the upper and lower flowers of each spikelet. The mean length of individually measured organs for the five spikelets was calculated. The logarithm of mean length for either the lodicule or stamen was plotted against the logarithm of mean glume length and a regression line with coefficient of correlation calculated for the wild type and ms-si using the graphics program "Sigma Plot".
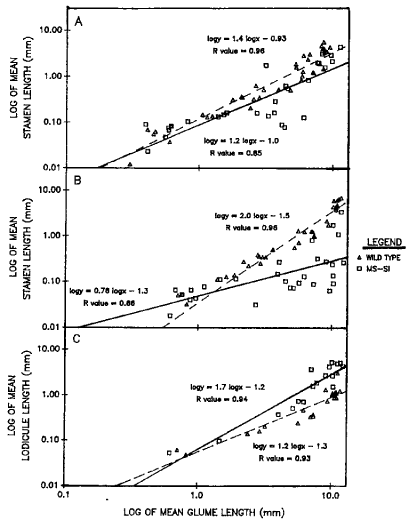
In Figure 1A, the slope of the regression line of upper floral stamen length measured against outer glume length for ms-si is slightly smaller compared to wild type. Although the differences between WT and ms-si may not be significantly different, additional comparisons of 1) stamen length to width measurements; 2) the length of silk-like structures, added to the plots, and 3) an examination for the development of pollen, may present significant differences as noted in our first contribution, this Newsletter.
Figure 1. Log mean lengths of outer glumes; stamens (excluding silk-like structures, SLS) from upper (A) and lower (B) and lodicules from upper (C) flowers of spikelets from the mid 1/3 portion of the main rachis of ms-si (ms-si/ms-si) and wild type (Ms-si/ms-si) tassels at various stages of maturity. The mean was calculated for five replicates or spikelets from the same tassel. Linear regressions are plotted to display the relationship between glume length and stamen or lodicule length. Differences in floral development between ms-si and wild type are depicted by the regression lines. R represents the coefficient of correlation.
The decrease in slope of the regression line of ms-si in Figure 1B reflects the ultimate abortion of the lower flower stamens in the ms-si spikelet. This developmental effect resembles in part the selective abortion of the lower flower in the ear.
In Figure 1C the enhanced slope of the regression line for ms-si upper floral lodicule measured against the outer glume depicts the development of lemma-like lodicules that are longer in total length than WT lodicules. Observations made of lower spikelet florets replicate the results noted, in this letter, of the upper flower. If measurements of length to width were to be plotted, differences in the two genotypes would also be shown.
Examination of spikelets from the tip one third and base one third of the main tassel rachis replicated the outcome noted for the mid-portion.
The important point to note from the plots is that floral organ differences between ms-si and WT can be detected at early stages of development in young spikelets. This observation further documents the statement, that ms-si is expressed early in floral development (MNL 63:88).
In conclusion, ms-si influences
floral development at the early stages, perhaps before or at the time of
organ primordia initiation. For ms-si genotypes, in comparison to
the wild type, the lower floral stamens abort and the lodicules are larger
in length.
Return to the MNL 64 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}