Pioneer Hi-Bred International, Inc.
SIOUX FALLS, SOUTH DAKOTA1
Augustana College
d13C = (R[sample]/R[standard] -1) x 1000
and R = 13CO2/12CO2.
This discrimination is greater among C3 plants, where the d value is often around -27� (parts/thousand), than among C4 plants, where it is often around -12�
It is now firmly established that variation in 13C discrimination in C3 plants depends on leaf intercellular carbon dioxide concentrations (Farquhar et al., Pl. Phys. 9:121, 1982; Farquhar et al., Annu. Rev. Pl. Phys., in press) and that the magnitude of the discrimination is related to water use efficiency and plant yield. This discrimination therefore holds promise as a surrogate for characteristics of importance in plant breeding. Selection strategies based on this measurement have been shown for wheat and barley (Hubick and Farquhar, Plant Cell Envir. 12:795, 1989), range grasses (Johnson et al., Crop Sci., in press), and other species. The factors which determine discrimination in C4 plants, for example maize, are more complicated (Farquhar, Aust. J. Pl. Phys. 10:205, 1983) and are less well documented. As an initial step in studying carbon discrimination in maize, we undertook a survey of selected cornbelt inbreds and hybrids to evaluate the range of discrimination in this germplasm pool.
Seeds were cleaned of cob material, ground to a fine powder and dried at 90 C. Approximately 1 mg samples were combusted in a Carlo Erba elemental analyzer, cryogenically purified for CO2, and analyzed in an isotope ratio mass spectrometer (SIRA 10). The precision of this analytical technique is <0.1�.
The results presented in Figure 1 show
that these samples exhibited a range of d13C values ranging from -11.65
to -10.74�. Although these values are quite positive for a C4 plant, they
are in agreement with a mean of -10.99� previously obtained from a collection
of CIMMYT cultivars. Those 33 cultivars were substantially more variable,
however, ranging between -12.00 and -9.76�. The smaller amount of variation
among the Pioneer samples is probably due to 1) the limited number of samples
analyzed
and 2) the fact that cornbelt germplasm
represents only part of the worldwide maize germplasm pool.
Figure 1. d13C values, in �, for seeds from 40 inbreds, 1 open pollinated variety (REID = Reid Yellow Dent) and 5 hybrids of maize grown under uniform conditions. Each point represents one analysis, except for the points shown as * which are the average of 3 analyses. These replicates agreed to within 0.2�. Two independently grown samples each of 3 inbreds were analyzed and are indicated in the figure.
Isotopic variation in maize should be determined by four main components: stomatal diffusion, hydration of CO2 in cell sap, PEP carboxylation of HCO3-, and loss of CO2 from the parenchymatous bundle sheath cells (Sasakawa et al., Pl. Phys. 90:582, 1989). Internal leaf CO2 (Ci) should change as the balance between diffusion and carboxylation varies and this change should be reflected in subsequent d13C measurements. Sasakawa et al. provided evidence that supports this relationship as they found increasingly negative d13C values as carboxylation capacity increased in maize leaves. Theoretically, increased diffusion resistance and potentially higher water use efficiency should also result in less negative values. However, there are as yet no experimental data to evaluate this relationship.
These data indicate that this sample
of cornbelt germplasm contains a relatively small fraction of the genetic
variation for d13C present in maize. This suggests a similarly small variation
in carboxylation activity and stomatal resistance in this germplasm. The
larger range of d13C values found in the CIMMYT samples, however, indicates
greater genetic variation in these physiological properties. This variation
may, in the future, be used to develop selection strategies that will aid
in the development of inbreds with improved water use efficiency.
Return to the MNL 64 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
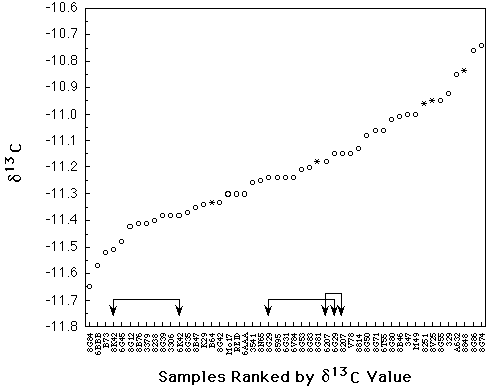{kind=link}