Siberian Inst. Plant Physiol. Biochem.
The mitochondria isolated from 3-day-old etiolated seedlings of Krasnodarsky 303 TV hybrid (W64A x Sg25) by the conventional method of differential centrifugation were used in the experiments. The mitochondrial pellet was resuspended in the solution containing 68 mM sucrose, 20 mM Na2HPO4, 4 mM KCl, 20 mM sodium succinate, 0.5 mM ATP, 10 mM MgCl2 and 50 mM Tris-HCl (pH 7.4). To translocate the plasmid DNA into mitochondria, 4-8 µg DNA of the pBR322 or pBR327 plasmid was added to the suspension (8-10 mg protein per 1 ml) and the mixture was incubated on ice bath for 30 min. The plasmid DNA that failed to bind to mitochondrial membrane was removed by thrice centrifugation of mitochondria. To protect mitochondria from possible contamination with nuclear DNA and to remove the plasmid DNA adsorbed on the surface of the mitochondrial membrane, the organelles were treated with pancreatic DNase (100 µg/ml) obtained from Serva (FRG). In the experiments on hybridization the electrotransfer of DNA from agarose gel to nitrocellulose membrane (Schleicher and Schuell) was performed in the Trans-Blot Cell apparatus (Bio-Rad) in tris-Borate buffer. The membrane was prehybridized in the standard solution at 65 C for 4 h and then hybridized with DNA of the pBR327 plasmid labelled with 32P GTP by means of nick-translation under the same temperature for 16 h. The nitrocellulose membrane was washed out and dried as described by Maniatis and exposed to X-ray film for some hours. The DNA was synthesized in mitochondria according to the method of Schegget and Borst (Biochim. Biophys. Acta 95:239-248, 1971) with the use of 32P ATP (specific radioactivity is 111 PBq mol-1). 32PNTP that failed to incorporate into the mitochondrial material was removed by centrifugation. To isolate and partially purify the mitochondrial DNA, the slightly modified method of Kemble et al. (Genetics 95:451-458, 1980) was followed. The DNA of pBR322 and pBR327 plasmids was purified by the method of alkaline extraction (Birnboim and Doly, Nucl. Acid. Res. 7:1513-1523, 1979) followed by gel filtration of the preparation on a column with sepharose CL-4B (Pharmacia, Sweden). Protein concentration in the mitochondrial fraction was determined by the Lowry method using bovine serum albumin as a standard. Electrophoresis of nucleic acids was performed in 0.8% agarose gel (Pharmacia). For radioautography the gel was dried and exposed to X-ray film for some days.
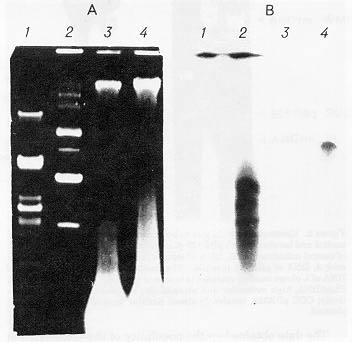
Electrophoretic and Southern blot analyses of DNA of maize mitochondria incubated with the pBR327 bacterial plasmid vector and followed by DNase treatment showed that under the conditions indicated the plasmid DNA was effectively translocated into the inner space of the organelles (Fig. 1A, B). It should be noted that only certain physical forms of the plasmid DNA were able to penetrate into the mitochondria. The illegible spot in the lower part of the gel in the radioautograph of the DNA from the treated mitochondria (Fig. 1B4) is likely to result from hybridization of the probe with degradation products of the plasmid DNA.
Figure 1. Electrophoretic (A) and Southern blot hybridization (B) analyses of DNA in maize mitochondria incubated with pBR327 plasmid. 1, PstI-fragments of l phage DNA used as markers; 2, pBR327 plasmid DNA; 3, DNA of control mitochondria; 4, DNA of mitochondria incubated with pBR327 plasmid. B, Radioautography from hybridization of blot A with pBR327 plasmid labelled with 32P in the reaction of nick-translation.
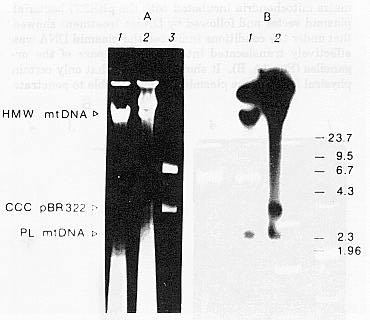
Figure 2 shows the results of radioautographic study of the DNA synthesis in isolated mitochondria incubated with DNA of the pBR322 bacterial plasmid. Several fractions of 32P-labelled DNA were registered to appear during incubation of the plasmid-treated mitochondria under conditions favourable for DNA synthesis in organello. Electrophoretic mobility of these fractions corresponded to the high molecular DNA of mitochondria, the covalently closed circular form of the pBR322 plasmid and the fraction of plasmid-like mitochondrial DNA. The use of the bacterial plasmid as a template for DNA synthesis by mitochondrial DNA polymerase in intact organelles is further evidence of the effective translocation of the plasmid DNA into the inner mitochondrial space. Moreover, only the covalently closed circular form of the plasmid was determined in the mitochondria which seemed to indicate the primary translocation of this molecular form or its preferential involvement into mitochondrial DNA synthesis as compared to other plasmid forms (open, circle and linear molecule). Nevertheless, further study is needed to clarify the nature of DNA synthesis of the pBR322 vector plasmid in maize seedling mitochondria (replicative or reparative).
Figure 2. Electrophoretic (A) and radioautographic (B) analyses of DNA of control and incubated with pBR322 plasmid maize mitochondria. 1, DNA of control mitochondria; 2, DNA of mitochondria incubated with the plasmid; 3, DNA of pBR322 plasmid. The position of HindIII-fragments of DNA of l phage used as markers is shown to the right. HMW mtDNA and PLmtDNA, high molecular and plasmid-like mitochondrial DNA, respectively; CCC pBR322, covalently closed circular form of bacterial pBR322 plasmid.
The data obtained on the possibility
of the translocation of bacterial plasmids into isolated mitochondria,
as well as the presence of the plasmid-like DNA set in the maize mitochondrial
genome (Kemble and Bedbrook, Nature 284:565, 1980), suggest the existence
of a specific mechanism of polynucleotide translocation in maize mitochondria.
Return to the MNL 64 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}