Dominant mutants at Kn1 (7 have been identified) are characterized by abnormal lateral vein development which results in protruding, hollow, finger-like pockets of tissue referred to as knots. Additionally, the Knotted mutation conditions ectopic and/or displaced formation of the ligule, a tissue which normally demarcates the leaf blade-sheath boundary. A specific, mild phenotype is associated with the Kn1-2F11 mutation. Typically, only the first one to three seedling leaves are affected by the formation of small knots, and ligule displacement is rare. The penetrance of the mutation varies and we have observed values from zero (no expression) to one (all individuals heterozygous for the mutation express the mutant phenotype). The expressivity of the mutation is also variable, but within the progeny of a given cross affected individuals usually express the mutation to similar degrees.
We suggested previously that variation in penetrance of Kn1-2F11 was correlated with the segregation of Ac or a gene closely linked to the Ac in our lines. The Ac element in our lines is on 9S, about 20 map units from Wx (K. Dawe, pers. comm.), and will be referred to as Ac-2F11. We have begun a genetic analysis to examine unlinked genes affecting Kn1-2F11, and the initial results of that analysis are reported here.
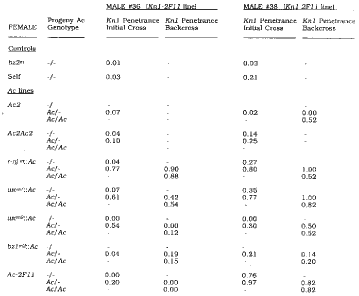
To test the effect of Ac on Kn1-2F11 expression, we used a two step crossing scheme. In the initial cross, an individual homozygous for the dominant mutation (Kn1-2F11/Kn1-2F11, ac/ac) was used as a male to many different Ac-containing females (kn/kn, Ac/Ac or Ac/ac). As controls, males used in the initial cross were selfed and crossed onto the bz2-m tester line. We used standard Ac elements located on 9S (wx-m7::Ac, wx-m9::Ac, bz1-m2::Ac, Ac-2F11) and 10L (r-njm::Ac), and Ac2 elements on 8L in one (referred to as "Ac2") or two closely linked (referred to as "Ac2Ac2") copies. We then selected progeny of the initial cross which were heterozygous for Ac and for Kn1-2F11 and backcrossed these individuals to the Ac line. The kernels from both crosses were separated by Ac dosage and planted to screen the progeny for the presence of knots. These data were used to calculate the penetrance (p) of Kn1-2F11 in each genotype.
The genetic analysis from progeny screens for two different males shows that Ac acts to increase the penetrance of the Kn1-2F11 mutation (Table 1). After crossing onto the no-Ac, bz2-m tester stock, both Kn1-2F11 lines show infrequent expression of the mutant phenotype. Line 36 shows knots only rarely after selfing (p=0.03), while the progeny of male 38 selfed exhibit higher penetrance (p=0.21). After crosses to Ac lines, however, penetrance is substantially higher in plus-Ac plants than in their no-Ac siblings. In general, the 38 line retains a higher "baseline" penetrance through additional crosses, and Ac acts to increase Kn1-2F11 penetrance above that baseline.
Table 1. Penetrance of Kn1-2F11 in progeny containing variable Ac dosage.
The results from the five standard Ac elements used in this study can be discussed in three groups according to penetrance scores from the initial cross. The Ac elements at r-njm::Ac and wx-m7::Ac both greatly increase the frequency of occurrence of knots. For the initial cross of each line, the penetrance in the plus-Ac seedlings is much higher than the baseline score in the no-Ac class (difference ranges from 0.42 to 0.73). In the backcross progeny, penetrance remains high and is similar for plants containing one or two doses of the element. As a second group, the Ac elements at wx-m9::Ac and Ac-2F11 are comparatively weaker in the initial crosses, as the penetrance difference between the plus-Ac and no-Ac classes ranges from 0.20 to 0.54. In the backcrosses to the wx-m9::Ac and Ac-2F11 lines, penetrance remains high or increases slightly in the presence of Ac for line 38. The line 36 backcrosses appear to uncover suppression, although the Ac-2F11 data may represent reversion of Kn1-2F11 by excision of Ds2. Finally, the crosses to the bz1-m2::Ac line give peculiar results. Contrary to the results seen with other Ac elements, none of the crosses in Table 1 show an increase in penetrance segregating with bz1-m2::Ac. Backcrosses to a homozygous Kn1-2F11, no-Ac line, however, show a striking increase in penetrance in the plus-Ac class over no-Ac siblings (difference = 0.4, data not shown). Either suppression has been relieved by two generations of crossing away from the bz1-m2::Ac line or the effect of this particular Ac element is only evident when Kn1-2F11 is homozygous.
The data also show that one and two doses of standard Ac in the plant affect penetrance similarly, but that the mutation is sensitive to Ac2 dosage. Ac2 is an anomalous Ac element that shows additive dosage on frequency of transposition events (Rhoades and Dempsey, MNL 57:14-17, 1983). Three (or more) doses of Ac2 are required to see bz2-m spots in the endosperm, while four doses are necessary to induce chromosome breaks at Ds. As indicated in Table 1, one dose of Ac2 has little or no effect on Kn1-2F11. In fact, in all initial crosses to the Ac2 line Kn1-2F11 showed low penetrance (p 2 0.07). In contrast, two doses of Ac2 in the plant increases penetrance of the mutation. This is seen in plants homozygous for Ac2 or heterozygous for a chromosome containing two closely linked Ac2 elements. The crosses to the Ac2 lines may uncover suppression, but in this case the enhancer effect of 2 doses of Ac2 confers expression of the mutant phenotype despite that suppression. Crosses are in progress to test for any combinatorial effects with Ac2 and standard Ac.
We are interested in the mechanism by which Ac influences expression of Kn1-2F11. Several points exclude somatic excision of Ds2 as the cause of knot formation. For example, plants carrying the mutation but lacking any active Ac elements are capable of expressing the mutant phenotype. In plants which do carry an active Ac element, individuals may be normal although Ds2 excision occurs, evident at both the bz2-m locus (plant and kernel sectors) and the Kn1-2F11 locus (Southern data). Furthermore, while Ac shows a dosage effect on the timing and frequency of Ds excision events, we see no dosage for standard-Ac enhancement of Kn1-2F11. Although Ds2 is inserted within transcribed sequences, we have not detected transcript differences between mutant and wild type at the level of Northern blots.
The crossing scheme we used defines
trans-acting factors influencing Kn1-2F11 expression. While the
data show that Ac is a dominant enhancer of Kn1-2F11, they
also suggest that some background suppression effects can preclude the
Ac effect. We are analyzing the background effects observed thus
far to determine whether any of them segregate as single genes. Furthermore,
we are investigating whether changes in the ability of Kn1-2F11
to respond to Ac are correlated with changes in methylation of the
Ds2 insertion at the Kn1-2F11 locus.
Return to the MNL 64 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}