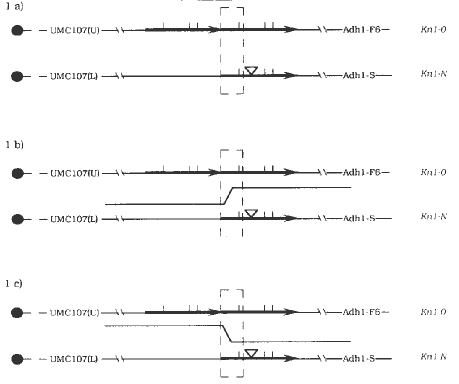
Plants that were doubly heterozygous for Kn1-N Adh1-S and Kn1-O Adh1-F6 (Figure 1a) were crossed to normal plants that were marked with a different Adh1 allele. The majority of the 2700 plants screened appeared to be either Kn1-O or Kn1-N heterozygotes. However, 2 plants were found that were normal and 3 plants were found that were extremely knotted. The normal plants were crossed and progeny obtained. We determined whether recombination had occurred in the derivatives with the flanking markers UMC107 and Adh1. UMC107 is proximal to Kn1 by 7 map units (Mathern and Hake, MNL 63:2, 1989); the polymorphism we have designated (U) is linked to Kn1-O, (L) is linked to Kn1-N. Adh1 is distal to Kn1 by 1 map unit. One normal derivative appears to be a contaminant by an assortment of markers. The other normal derivative does not carry the tandem duplication of Kn1-O nor the insertion of Kn1-N. It carries Adh1-F6 and UMC107(L). Therefore, it is likely to have arisen following unequal crossing over in Figure 1b. The knotted plants were too severe to obtain a cross, but DNA was isolated and analyzed. The DNA of one of the severe plants appears similar to Kn1-N and may be due to an unlinked gene increasing the severity of Kn1-N or a small DNA alteration at Kn1-N. The DNA from the other 2 severely knotted plants appears similar to both Kn1-O and to Kn1-N. The tandem duplication characteristic of Kn1-O is present as well as the insertion that has come to characterize the Kn1-N mutation. Both derivatives carry the proximal marker of the Kn1-O chromosome, UMC107(U), and the distal marker of Kn1-N, Adh1-S. Thus it appears, as shown in Figure 1c, that recombination has occurred, replacing the distal repeat of Kn1-O with similar DNA from Kn1-N.
Figure 1. Intragenic recombination at knotted. Chromosome constitution of a) heterozygous progenitors, b) possible recombination event to produce normal derivative, c) possible recombination event to produce severe Knotted derivative. Dashed box delimits recombination region.
We do not know the cause of the Kn1-N
mutation as we do with Kn1-O or Kn1-2F11, which result from
a tandem duplication and an insertion of Ds2, respectively. The
results of this recombination delimit the lesion that causes Kn1-N
to the right of the recombination event in Figure 1c. It suggests that
the 500 bp insertion may be the cause of the Kn1-N mutation but
with such few numbers and without the reciprocal recombinant, it is impossible
to say. Similar to the Bar locus in Drosophila, the Kn1-O
locus undergoes reversion at a high rate (1 in 3000). We have isolated
derivatives from homozygous Kn1-O plants that have gained an extra
copy of the tandem duplication and are more severely knotted, and have
a number of derivatives that have lost the duplication and are normal.
The rate of recombination with the Kn1-O/Kn1-N double heterozygotes
is equally high, if not higher.
Return to the MNL 64 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}