Plant Gene Expression Center, USDA-ARS
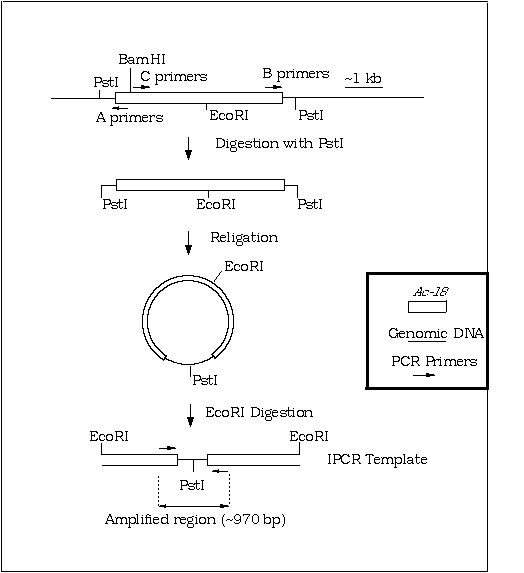
Figure A. Schematic representation of IPCR template generation from genomic tobacco DNA carrying Ac-18 inserted at the Nt-1 locus. For amplification of sequences flanking both ends of the element, genomic DNA was digested with PstI, ligated at a DNA concentration of 2 µg/ml and then linearised with EcoRI to yield the illustrated template. Primers not to scale.
The number of enzymes suitable for the initial cut in the above IPCR scheme is restricted to those which do not cleave within the Ac element and is further reduced to a sub-group of these with sites in the flanking genomic DNA relatively close to each end of the inserted element. In general application, where often the goal is to obtain a probe for the target region, amplification of sequences flanking only one end is sufficient. This enables the utilization of the many frequent-cutting enzymes with sites within the element for the initial digestion. This notion was used to generate 'short-cut' IPCR templates from the Nt-1::Ac-18 locus. Primers from locations A and C were utilised in this procedure. Template was produced by initial digestion with enzymes cleaving within the genomic DNA and 3' of the C primers within the element followed by religation and linearization with BamHI. Intermolecular ligation in the template generation protocol, producing various chimeric templates, was found to be a potential problem when frequent cutting enzymes were utilized for the initial digestion (the frequency of intermolecular ligation increasing with the concentration of ligatable ends). This was overcome by the use of a methylation sensitive enzyme (EcoRII) for the first cleavage (which resulted in frequent digestion within and around the element while leaving much of the genomic DNA in relatively large pieces).
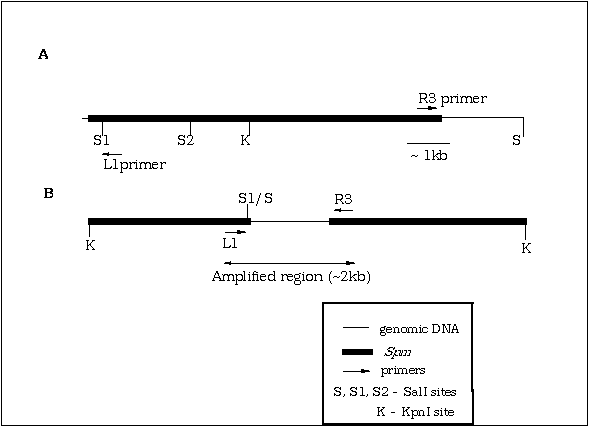
Subsequently we utilized this methodology to amplify and clone sequences flanking an Spm element in a line of maize derived from a c2-m1 line. A specific Spm insertion, which was characterized as a 10.2 kb SalI fragment hybridizing to an Spm derived probe, was selected as a target for amplification. Primers were designed to prime synthesis out of the 5' end of the element (primer L1, Figure B) from the region immediately 3' of the SalI site at position 276 (this SalI site is unmethylated in active elements, Banks et al., Genes Dev. 2, 1364-1380, 1988) and from the 3' end of the element (primer R3). Genomic DNA was size fractionated to enrich for this 10.2 kb SalI fragment, and IPCR template generated as illustrated in Figure B. Amplification yielded a product of ca. 2 kb which hybridised to the 10.2 kb band and to an additional band of 2.0 kb in lines heterozygous for the Spm insertion. The IPCR product was used as a probe to map the location of this element to position 112 on chromosome 4L (B. Burr, personal communication).
Figure B. Schematic representation of IPCR template generated from genomic maize DNA.
(A) Partial restriction map of Spm showing locations of PCR primers. The genomic SalI site 3' of the Spm element selected as a target for IPCR is also indicated.
(B) IPCR template derived from the selected
Spm shown in (A) by ligation of SalI digested size fractionated
DNA and subsequent linearisation with KpnI. Primers not to scale.
The method should provide considerable
advantages in obtaining element flanking sequences compared with conventional
lambda cloning and screening. IPCR amplification of genomic sequences flanking
transposed elements will facilitate the subsequent isolation of wild type
genes in tagging experiments and will also generate RFLP probes for use
in genome mapping.
Return to the MNL 64 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}