Montana State University
MADISON, WISCONSIN
University of Wisconsin
--Victor Raboy and Oliver E. Nelson Jr.
The bz-m13 allele of the bronze-1 (bz) locus contains a 2.2kb defective Suppressor-mutator (dSpm) insertion in the second exon of bz. In the presence of an active, standard Spm (Spm-s), the dSpm insertion in bz-m13 is excised early during development, resulting in 1) large revertant sectors in the aleurone and 2) high rates of germinal excision. Six change-in-state (CS) derivatives of bz-m13 have been isolated and studied in some detail (Schiefelbein et al., PNAS 82:4783, 1985). Their dSpm inserts, in response to Spm-s, are excised later in development, resulting in characteristically small revertant sectors (spots) in the aleurone and few if any germinal excisions. A molecular analysis revealed that five of the six CS's dSpms were deletion derivatives of bz-m13's dSpm. The sixth CS, bz-m13CS1 (CS1), is identical in sequence to bz-m13. The delay in timing of dSpm excision characteristic of this CS is attributable to the fact that certain sequences within CS1's dSpm are highly modified, whereas they are not in bz-m13. The DNA modification probably consists of methylation of cytosine in CG and CXG sequences. The sequence-specific DNA modification we observe and its relationship to the activities of bz-m13 and CS1 is very similar to that found between active and inactive Spm elements as described by Banks and Fedoroff (MNL 62:27).
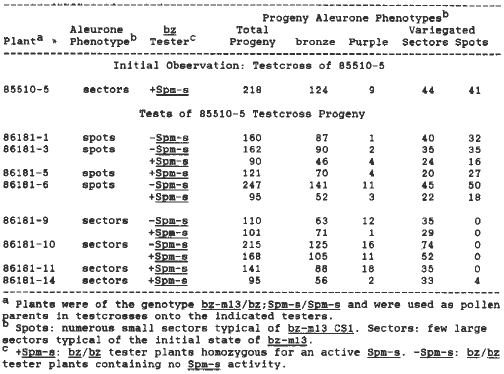
To test the correlation between DNA modification and the CS1 phenotype we sought to isolate a number of novel "CS1-like" derivatives. Plants of the genotype bz-m13/bz; Spm-s/Spm-s were crossed by and on tester stocks of the genotype bz/bz; Spm-s/Spm-s. New isolates appeared either as 1) one or a few individual kernels on a testcross ear, representing events which occurred relatively late during development, or as 2) the result of an event which occurred relatively early during development, so that all male and female gametes receiving the bz-m13 allele contained CS1-like derivatives. In 1985, five of 82 plants tested gave this latter result. In one exceptional case, the testcross of 85510-5, approximately half of the variegated kernels displayed a bz-m13-like aleurone phenotype (few large sectors), and half displayed a CS1-like phenotype (numerous small sectors) (Table 1). How is it that when we testcross a plant of the genotype bz-m13/bz, we observe two variegated phenotypes in approximately equal numbers?
Kernels representing each of these two classes were grown out in 1986 and crossed onto bz/bz tester plants that were either homozygous for Spm-s (+Spm-s) or lacking in Spm-s activity (-Spm-s) (Table 1). It is relatively easy to distinguish the difference in phenotype between bz-m13 and CS1 when the endosperm contains one copy of a given allele, but difficult to distinguish when it contains two copies. Regardless of tester, plants grown from kernels that were CS1-like (86181-3, -5, -6) reproduced the initial result observed with 85510-5. With plants grown from kernels that were bz-m13-like (86181-9, -10, 11, -14), essentially all progeny receiving the bz-m13 allele were bz-m13-like.
Table 1. Testcross analyses of the initial exceptional plant 85510-5 and its variegated progeny.
Our first hypothesis was that either an altered Spm element such as an Spm-weak, or a dominant modifier of Spm-s, was segregating independently of bz, and was responsible for this unusual pattern of inheritance. We know that bz-m13, in response to an Spm-w or En-low, is excised late during development, mimicking the CS1 phenotype. We therefore tested a total of 34 bz kernels from the testcrosses of 85510-5, 86183-3 and 86181-6 for the presence of a modified Spm or a dominant modifier of Spm-s. The results indicate that these plants were homozygous for Spm-s. No altered Spm or modifier of Spm was found. Thus the phenomenon responsible for this exceptional pattern of inheritance appears to be specific to the bz locus.
Our current hypothesis is that this phenomenon is an outcome of strand-specific, asymmetric DNA methylation. Such a phenomenon has been proposed as responsible for the developmental asymmetry of daughter cells in fission yeast (Schizosaccharomyces pombe) (Klar, Nature 326:466, 1987). Perhaps the interaction of Spm-encoded products and DNA methylases results in this strand-specific modification. The separation of chromatids following the second division of meiosis might then produce two daughter cells which contain the same bz-m13 sequence, but which represent different patterns of methylation, which in turn results in the different phenotypes.
A second possibility is that the modification event occurs frequently
following the second microspore division, resulting in a high rate of noncorrespondence
in DNA methylation pattern between germ and endosperm, coincidentally approximating
1/2 of bz-m13 progeny. Subsequent tests of 86181-3 and 86181-6 progeny
have not ruled out this possibility. CS1-like kernels from testcrosses
of these two plants mostly produce all CS1-like progeny, and occasionally
reproduce the results observe with 85510-5. bz-m13-like kernels
from these two plants often produce only bz-m13-like progeny, but
occasionally produce CS1-like progeny and combinations of the two.
Whatever the mechanism, this phenomenon is not unique to 85510-5 and its
progeny. We have identified several additional independent cases.
Return to the MNL 63 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}