Carnegie Institution of Washington
--P. Masson, G. Rutherford and N. Fedoroff
The Spm family of maize transposable elements consists of both autonomous and defective elements. The former are capable of promoting their own transposition, as well as the transposition of the defective elements (dSpm elements). The latter are totally stable in the absence of a trans-acting autonomous Spm element in the same genome and are deletion derivatives of the autonomous element (reviewed by Fedoroff, in Howe, M. and Berg, D., eds., "Mobile DNA", in press).
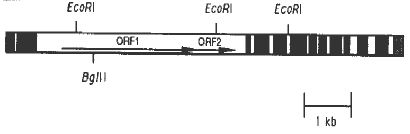
The wild-type autonomous Spm-s element is 8.3kb long; its structure is shown in Figure 1 (Pereira et al., EMBO J. 5:835-841, 1986, Masson et al., Genetics 177:117-137, 1987). Between the 13bp terminal inverted repeats and the 250-300bp subterminal regions, the Spm-s contains two long open reading frames (ORF1 and ORF2). Element transcription yields one major 2.5kb transcript, whose structure is also represented in Figure 1, and two minor 6kb transcripts (Pereira et al., 1986, Masson et al., 1987, Banks et al., Genes & Develop., 2:1364-1380, 1988). The 6kb transcripts overlap the two ORFs and the right end of the element.
Figure 1. Structure of the Spm-s element. The two 13bp inverted repeats and 250-300bp subterminal regions are represented by the stippled boxes on each end of the element. The sequences encoding the exons of the major 2.5kb transcript are represented by black boxes. The two long open reading frames are represented by arrows. The EcoRI and BglII sites are also indicated (Pereira et al., EMBO J. 5:835-841; Masson et al., Genetics 177:117-137).
Despite a good knowledge of the Spm structure, very little is known about the actual Spm sequences encoding the transposase. To gain more insight into this function, we have tested the transposition of Spm in tobacco. The Spm-s element was introduced into tobacco by Agrobacterium-mediated transformation, and its transposition was analysed by the detection of flanking plasmid sequences lacking Spm sequences (empty donor fragments) in the DNA extracted from transgenic plants (Masson et al., MNL 62: 26-27, 1988). We have cloned and sequenced fifteen empty donor sites from one transgenic plant (Table 1). The empty donor site fragments can be classified into three different categories, deriving from either precise excision or imprecise excision of the Spm element. All sequences are among the types of empty donor fragment sequences observed after Spm transposition in maize (Schwarz-Sommer et al., EMBO J. 4: 591-597). This observation, coupled with the detection of transposed copies of Spm in the DNA extracted from the same transgenic tobacco plants, demonstrates that Spm transposes in tobacco as it does in maize (Masson and Fedoroff, PNAS, in press).
Table 1. The nucleotide sequences of empty donor sites.
| Original sequence: | ||
| ...TTTGAATATT-Spm-AATGGA... | ||
| Empty donor fragment sequences: | ||
| -Group 1: precise excision of Spm | ||
| ...TTTGAATATT AATGGA... (2 sequences)* | ||
| -Group 2: imprecise excision of Spm | ||
| ...TTTGAATATT ATGGA... (6 sequences)* | ||
| -Group 3: imprecise excision of Spm | ||
| ...TTTGAATAT TTAATGGA... (7 sequences)* | ||
*For each group of empty donor fragments, the number of cloned fragments having the sequence shown is indicated in parentheses.
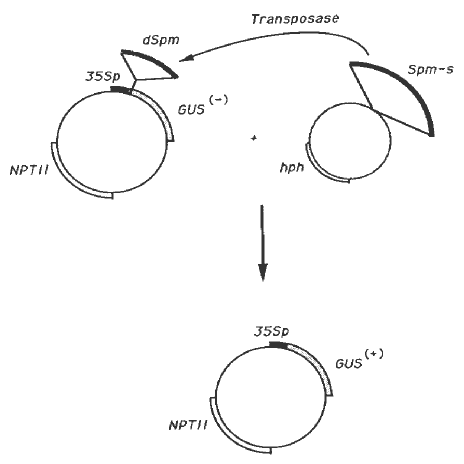
Figure 2. Schematic representation of the assay for Spm transposition in tobacco. The Spm-s element, inserted in a plasmid conferring hygromycin resistance (hph gene) to the plant cell, provides the transposase function necessary to induce the excision of dSpm inserted within the GUS gene on a plasmid conferring kanamycin resistance (NPTII gene). 35Sp refers to the 35S promoter from CaMV (Shah et al., Science 233:478-481). (+) and (-) refer to the activity of the GUS gene. The vectors were generously provided by Dr. S. Rogers (Monsanto Co.).
To study the Spm sequences encoding the transposase function, we have developed a functional assay which allows us to test the ability of in vitro-mutagenized Spm elements to promote the transposition of a defective element in tobacco (Masson and Fedoroff, PNAS, in press). The assay is based on the restoration of GUS gene expression after dSpm excision (Figure 2). The E. coli GUS gene, coding for ß-glucuronidase, is functional in plants when fused to the appropriate control sequences (Jefferson et al., EMBO J. 6: 3901-3907). The ß-glucuronidase enzyme converts the chromogenic X-GLU substrate into an insoluble blue compound which accumulates in the transformed plant cells expressing the GUS gene. A dSpm element inserted between the promoter and the open reading frame of the GUS gene disrupts GUS gene expression. However, when the same plasmid is introduced in tobacco together with a second plasmid carrying a wild-type Spm-s element, the latter induces dSpm excision. This results in the appearance of cells expressing the GUS gene (Figure 1). During development, such cells divide and generate sectors of X-GLU-stainable, GUS-positive cells, whose number and size reflect, respectively, the frequency and timing of transposition.
Using this assay, we have shown that Spm-s induces the excision
of a defective dSpm element introduced on a separate Ti plasmid.
Spm transposition occurs late and infrequently in tobacco. However,
earlier transposition often occurs when the trans-acting Spm-s element
is expressed from the CaMV 35S promoter (Masson and Fedoroff, PNAS, in
press). In vitro -mutagenized Spm elements are being
tested for their ability to promote the excision of a standard dSpm
element inserted in the GUS gene. Preliminary data indicate that ORF1 and
ORF2 are necessary for the transposase function of Spm: frameshift
mutations in the EcoRI and BglII sites in ORF1, or in the
EcoRI site in ORF2 (see Figure 1 and Table 2) alter the transposase
function of the element. On the other hand, a frameshift mutation in the
EcoRI site located in the intron sequence separating the fourth
and the fifth exons of the major 2.5kb transcript (Figure 1) does not affect
this function (Table 2). However, our previous analysis of several Spm
derivatives in the a-m2 alleles of maize indicates that the right
end of the element, including sequences encoding the last ten exons of
the major 2.5kb transcript, is also necessary for the transposase function
of the element (Masson et al., 1987).
Table 2. Transposase function of in vitro-mutagenized Spm
elements in tobacco.
| Site of frameshift mutation (1) | Number of calli analysed | Number of calli showing GUS-positive sectors (2) |
| ORF1: | ||
| EcoRI | 14 | 1# |
| BglII | 14 | 0* |
| ORF2: | ||
| EcoRI | 9 | 0 |
| 4th intron: | ||
| EcoRI | 20 | 9# |
| Wild-type: | 12 | 10# |
| Defective (3): | 25 | 0 |
(1) The frameshift mutagenesis was done by cutting the element with
the appropriate restriction enzyme (see Figure 1 for their position on
the Spm sequence), filling in the ends using the Klenow fragment
of the E. coli polymerase and religation (Maniatis et al.,
In "Molecular Cloning: A Laboratory Manual", CSHL).
(2) The in vitro-mutagenized Spm elements were tested
for their ability to promote excision of dSpm in tobacco. dSpm
excision was monitored by the detection of X-GLU-stainable, GUS-positive
sectors, as described in the text. # The positive calli showed multiple
GUS-positive sectors. * Two calli showed one single GUS-positive sector.
(3)In the negative control experiment, the reporter plasmid containing
the dSpm element inserted within the GUS gene was co-transformed
in tobacco together with the second vector which contained no Spm
element.
Taken together, the data presented here indicate that ORF1, ORF2 and
sequences in the right half of the Spm element are necessary for
the transposase function of the element. These various sequences are present,
at least in part, in the minor 6kb transcripts encoded by the element.
We have recently cloned partial cDNAs corresponding to internal portions
of the 6kb transcripts produced in maize. Preliminary results indicate
that several RNA species are produced by alternative splicing between sequences
in ORF1, ORF2 and the sequences encoding the translated exons of the major
2.5kb transcript (not shown). We are currently cloning the remaining portions
of the cDNAs for the 6kb transcripts.
Return to the MNL 63 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}