--N. M. Antonelli and J. Stadler
A method for transfection of Black Mexican Sweet protoplasts (line BMS-M) with polyethylene glycol was adapted from Krens (Krens et al., Nature 296:72, 1982) and reported in this Newsletter (MNL 62:7,1988). In that article we described the detection of transient expression of transfected chloramphenicol acetyl transferase (CAT) genes. We now report the recovery of stable kanamycin-resistant microcalli obtained after polyethylene glycol (PEG)-induced transfection of BMS-M protoplasts with the neomycin phosphotransferase II gene (pCaMVNeo; Fromm et al., 1986). This frequency of stable transformation of maize protoplasts is higher than that reported by others (Fromm et al., Nature 319:791, 1986) and may reflect the somewhat higher average viability of protoplasts 24h after PEG treatment (70%) compared to electroporated protoplasts (50%).
To obtain stable transformants we used two methods of protoplast culture after PEG-mediated gene transfer. In the first procedure the protoplasts were resuspended immediately after treatment in growth medium containing 8% mannitol and 25% conditioned medium (CM)(Somers et al., Plant Sci. 53:249, 1988) and incubated in the dark at 28 C for 14 d. Then, to allow more cell division under nonselective conditions, 1ml aliquots of cells in liquid were placed on solid growth medium (containing 0.3% Gelrite and 25% CM) for an additional 7d. After this time 150 individual microcalli were picked at random and transferred to selective solid medium containing either 100 or 200ug/ml kanamycin and 25% CM, and were incubated further for 7 to 14 d. At the end of the second period on selective medium, some microcalli were faster growing than others (Fig. 1), and these were harvested for molecular analysis when they were approximately 10mm in diameter with 300-400mg tissue. In the experiment presented in Table 1, 31 of the 150 microcalli picked were fast growers. These microcalli, however, may not have been of unique clonal origin since treated protoplasts tend to aggregate in liquid culture after PEG treatment, and during growth in liquid for 14d the larger aggregates often subdivide to form smaller (possibly genetically identical) clusters. There was either no growth in selective conditions of microcalli from control cultures that had received PEG treatment but no DNA or a limited amount of cell division followed by browning of the callus in a few weeks. Genomic DNA from the faster growing microcalli was isolated (Mettler, Plant Mol. Biol. Rep. 5:346, 1987) and digested with several restriction enzymes including HindIII and BamH1. A probe containing the BamH1 fragment of the nptII gene was radioactively labeled and hybridized. The 1.0kb diagnostic fragment demonstrating nptII genomic insertion was present in 5 of the 31 faster- growing colonies (Table 1; Southern blot not shown).
Fig. 1. Selection for stable transformant colonies. The arrow indicates some fast-growing calli on 200ug/ml kanamycin.
Table 1. Selection of kanamycin-resistant cell lines after treatment
of BMS protoplasts with PEG and pCaMVNeo (50ug.)
| a | b | c | d | e | |
| Control | 5x106 | 2x106 | 50 | 0 | 0 |
| Liquid | 5x106 | 2x106 | 150 | 5 | 3.3 |
| Agarose slabs | 5x106 | 2x106 | 150 | 5 | 3.3 |
a. Number of treated cells.
b. Cell number after PEG treatment.
c. Total number of colonies picked at random.
d. Transformed colonies from population identified by fast growth in
selective conditions.
e. Transformation frequency (% of randomly recovered colonies).
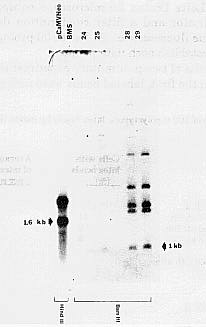
In the second procedure, protoplasts were plated immediately after PEG treatment in a 2ml slab of 0.6% low melting agarose (Paszty and Lurquin, BioTechniques 5:716, 1987) in growth medium with 6% mannitol and 25% CM which was then overlaid with less than 0.5ml liquid growth medium also containing 6% mannitol and 25% CM. After 2 weeks, the slabs were sectioned and placed as above on solid nonselective growth medium for additional proliferation of cells for 7d before selection. Rapid growth of discrete microclusters was observed during this second round of growth on nonselective agar. These microclusters are called "colonies" since the original aggregates of PEG- treated protoplasts were maintained as unique entities embedded in agarose. This plating method does not allow further division into sib aggregates during growth on nonselective medium. Further, it is also possible that each original cluster of PEG-treated cells may have contained no more than one transfected cell. A random sample of the colonies arising was transferred to the described kanamycin-containing selective medium, and after 7d the faster- growing transfected microcalli were harvested for Southern blot analysis. There was again little or no growth of colonies originally derived from PEG- treated cells without DNA. Five of the 7 fast-growing colonies picked showed the 1.0kb band expected after hybridization of genomic DNA containing integrated nptII genes with the BamH1 nptII probe (Fig. 2). Other bands of larger size in Fig. 2 are the result of probe contamination with radioactively labeled residues of the vector (pUCPiAN7) that contains the nptII gene in pCaMVNeo.
The common feature shared by these two methods for recovery of transformants is the extended time for proliferation of treated cells in nonselective medium. When in both instances a random population of healthy, actively growing microcalli or colonies was later transferred to kanamycin-containing medium, the selective medium allowed identification after 7d of a subpopulation of fast growers. In the experiments presented (Table 1), 31 of the 150 microcalli isolated from populations of treated cells initially grown in liquid medium were fast growing, as were 7 of the 150 colonies picked from the population plated in agarose slabs. Rapid growth on kanamycin identified transformed cells exhibiting stable drug resistance since 16% to 71% of these fast growers were proved transgenic by molecular analysis. The overall frequency of transgenic isolates was at least 3.3% of randomly selected microcalli and may be higher since some slower growing microcalli may also have been transformed. This frequency of transformation of corn cells is greater than reported by others (Fromm et al. 1986). We have established cell lines from the microcalli that were positive for nptII gene insertion, and these continue to grow briskly on medium containing 200ug/ml kanamycin.
Fig.
2. Southern blot analysis of BMS-M kanamycin-resistant calli. The pCaMVNeo
was digested by HindIII, generating a 1.6kb fragment containing
the nptII gene (on the left side). The probe used was the 1.0 kb BamHI
fragment containing the nptII gene. No hybridization occurs in BMS-M control
DNA. Samples 24, 25, 28, 29 show the diagnostic 1.0kb fragment as well
as larger bands which are due to probe contamination with residues of pUCPiAN7.
Rehybridization of the same filter with a labeled 3.4kb BamHI fragment
of pCaMVNeo gave the same pattern minus the 1.0kb fragment (data not shown).
Return to the MNL 63 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}