--J. R. Laughnan, S. Gabay-Laughnan and Janet M. Day
A number of inbred lines of maize carry a nuclear gene capable of restoring male fertility in plants carrying S male-sterile cytoplasm (cms-S). The cms-S restorer gene designated Rf3 exhibits a gametophytic mode of restoration such that Rf3 rf heterozygotes produce 50 percent Rf3-carrying pollen grains that function normally in fertilization, and 50 percent rf-carrying grains that abort. Tests of allelism of the Rf3 gene carried by ten inbred lines, including Ky21, CE1 and Tr, indicated that all are allelic (see Maize Breeding and Genetics, 427-446, 1978). Some of these tests involved crosses between two cms-S restoring inbred lines followed by testcrosses of the F1 plants with (S) rf rf male-sterile testers; with this protocol the absence of male-sterile plants in testcross progenies indicated either allelism or very close linkage of the cms-S restorers carried by the parental inbred lines. Another procedure involved crosses of a cms-S restoring inbred line as female parent with another restoring line carrying cms-S or normal cytoplasm; since the pollen of F1 plants was all normal, where 25 percent of grains of the shriveled type characteristic of abortion found in the cms-S system is expected if the restorers are nonallelic and segregate independently, it was concluded that the restorer genes carried by the F1 plants are either allelic or very closely linked. Another procedure, involving testcrosses of F1 plants heterozygous for an inbred-line restorer gene, Inversion 2a and the B allele, sufficed to assign the cms-S restorer(s) involved to chromosome 2, but was not useful for determination of allelism of inbred line restorers or for mapping their positions in chromosome 2.
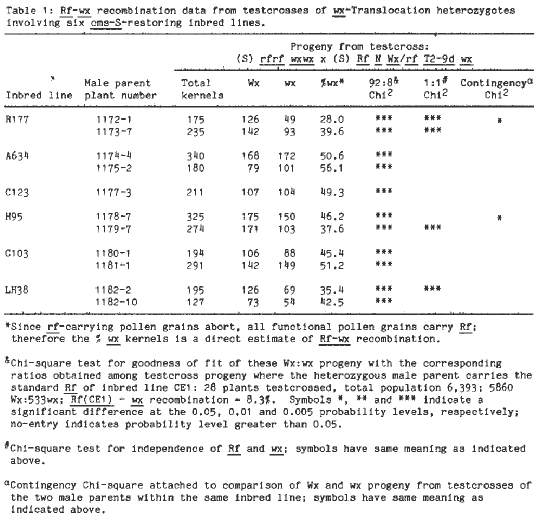
In the final analysis the chromosomal location of Rf3 in inbred lines CE1 and Tr was determined by testcrosses of cms-S wx-translocation heterozygotes, that were also heterozygous for an inbred line cms-S restorer, onto wx wx testers. For male parents carrying the CE1 restorer the testcross may be illustrated: (S) rf rf wx wx X (S) Rf(CE1) N Wx/rf T2-9d wx. Since pollen grains carrying the nonrestoring allele abort, only Rf(CE1) grains function so that the percentage of waxy kernels on these testcross ears provides a direct estimate of Rf(CE1)-wx recombination. We have extensive data on Rf(CE1)-wx recombination in T2-9d (breakpoint 2L 0.8) heterozygotes. Some of these are given in the second footnote of Table 1, where this recombination rate, with over 6000 kernels scored, is 8.3 percent. The cms-S restorer carried by inbred line Tr has most often given similar recombination rates in testcrosses involving wx-T2-9d heterozygotes, but at least one subline of Tr inbred line has an Rf(Tr)-wx recombination of 17 percent with a total of 3018 kernels scored.
Because of the apparent discrepancy indicated above we commenced experiments
to determine Rf-wx recombination rates in T2-9d heterozygotes that
carry cms-S restorers from a number of inbred lines. The results of testcrosses
involving six such lines are given in Table
1. Since the crosses that produced the male parents here involved a
cms-S inbred line as female parent and a highly inbred strain of homozygous
wx-T2-9d as male parent, the testcrossed sibling F1 plants were
isogenic. Surprisingly, the Rf-wx recombination rates for all six
inbred line sources showed highly significant departure (at 0.005 level)
from the Rf(CE1)-wx rate (8.3%). In fact, the recombination rates
for restorers carried by inbred lines A634, C123 and C103 do not differ
significantly from a theoretical 1:1 ratio, indicating that these restorers
assort independently of the chromosome 2 wx-linked allele in the
T2-9d heterozygote and may be presumed either not to be in chromosome 2
or to be located in chromosome 2 at some considerable distance from the
wx locus in the translocation heterozygote. In both H95 and LH38,
one tested sibling in each shows a highly significant departure from a
1:1 ratio, while the other is in good agreement with it. Since these F1
siblings are, in all other regards, presumed to be isogenic, these differences
in Rf-wx recombination rates for sibling F1 plants indicate that
Rf(H95) and Rf(LH38) can occupy different chromosomal sites
in sibling plants within each inbred line. These results are most easily
explained if it is assumed that these restorer elements have transposed
in recent or not so recent generations. Note that the Rf-wx recombination
rates for the two sibling H95 F1 plants show a significant difference in
the contingency Chi-square analysis and that this further supports the
transposition hypothesis. Both sibling parents carrying the R177 restoring
allele, as well as one of the sibling plants from each of the inbreds H95
and LH38, exhibit recombination rates that differ significantly from the
Rf(CE1)-wx rate as well as from a theoretical 1:1 expectation. The
recombination rates (28.0%, 39.6%, 37.6%, and 35.4%) indicate Rf
chromosomal sites linked with the Rf(CE1) site in chromosome 2.
The contingency Chi-square analysis for the R177 siblings indicates a significant
difference in Rf(R177)-wx recombination rates, suggesting that these
siblings carry the restorer at different sites in chromosome 2. In view
of the various chromosomal sites for cms-S restorers, the possibility that
any one inbred line may carry two or more restorer genes should not be
ignored. Although there is presently no evidence for this, it would explain
why the test for allelism of Rf genes in two different cms-S restoring
inbred lines would be positive even though these two lines might show significantly
different Rf wx recombination rates in the wx-T2-9d testcross
system; both lines could carry an Rf gene at the same site, e.g.
Rf(CE1), thus providing a positive test for allelism, but one of
the lines might carry a second restorer at a site some distance from the
first in chromosome 2 which, it can easily be shown, would increase the
frequency of Rf-wx recombinant kernels on testcross ears. Given
such a wx-T2-9d heterozygote in which the two Rf genes are
at maximum distance the frequency of waxy kernels on testcross ears approaches
a maximum of 33%, and the same is true if the second restorer is located
in a chromosome other than 2. This means that in this testcross system
a Wx:wx ratio of 1:1 indicates the presence of only 1 cms-S
restorer at a site in chromosome 2 or elsewhere such that it shows independent
assortment with wx in the wx-T2-9d testcross heterozygote.
Return to the MNL 63 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}