--G. Zabala, S. Gabay-Laughnan and J. R. Laughnan
The first studies of mitochondrial DNA (mtDNA) of male-fertile revertants from S-type male-sterile (cms-S) plants suggested the involvement of two episome-like double-stranded linear DNA molecules, S1 and S2 (Science 209:1021-1023, 1980; Molec. Appl. Genet. 2:161-171, 1983). Upon reversion, these free S-episomes disappear from the mitochondria and rearrangements involving S2 or S1 homologous sequences take place in the main mitochondrial genome. These results were consistent in revertants of cms-S plants with an M825 or 38-11 nuclear background. In a WF9 nuclear background however, cms-S revertants maintain the free S-episomes. Their homologous sequences in the main mitochondrial genome do not undergo rearrangement (Theor. Appl. Genet. 75:659-667,1988) but the s-R and y-R integrated sequences that are targets for recombination with the free S-episomes are missing (Theor. Appl. Genet. 76:609-618, 1988). These results indicate that the nucleus has its influence on mtDNA organization either during or prior to reversion to fertility.
The influence of the nuclear background on mtDNA has been demonstrated further by the conversion of cms-S lines having nuclear backgrounds WF9 and 38-11 to the nuclear background of M825 by backcrossing with M825 as the recurrent male parent. In revertants recovered subsequent to these nuclear conversions, the pattern of reorganization of the mitochondrial genome follows the pattern unique to revertants with the M825 nuclear background. All revertants lose the free S1 and S2 episomes and the S2 sequences integrated in the main mitochondrial genome suffer disruption of the left unique end (Escote, Ph. D. Thesis U. of Ill).
Nuclear action on mtDNA organization is not exclusive to the reversion event but is also observed in the absence of reversion. The sterile F1 progeny from the cross RD-WF9 x M825 (RD is a subtype of S) have an mtDNA organization different from that of the original RD-WF9 female parent. Differences are detected as the loss, starting as early as the F1 generation, of some S1, S2 and cytochrome oxidase subunit I (COXI) DNA fragments present in the parental RD-WF9. In the progeny of the first backcross of RD-WF9 with M825, additional RD-WF9 parental bands disappear, after which the Southern blot pattern is stable through the fourth backcross (Escote et al. MNL 60:127).
To determine whether the change in mtDNA organization already observed in the sterile F1 hybrids has any effect on the mtDNA rearrangements which occur upon reversion, we have analyzed the rearrangements in newly arisen cytoplasmic revertants of an F1 generation resulting from a cross between RD-WF9 and M825. The F1 hybrid plants have the mitochondria of the female parent, cms-RD, and a hybrid nucleus: 50% WF9 and 50% M825.
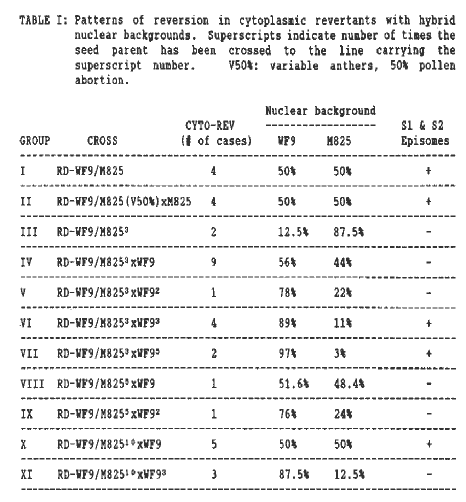
In Table 1 we have listed the eleven groups of cytoplasmic revertants (cyto-revs) used for this study. Five of them (I,II,IV,VIII,X) were recovered from plants carrying hybrid nuclei of approximately 50% WF9 and 50% M825 and six groups (III,V,VI,VII,IX and XI) derived from plants with differing WF9:M825 nuclear dosages.
When undigested mtDNAs were analyzed (Table 1), the following patterns emerged: cyto-revs from groups I,II,VI,VII and X maintained the S1 and S2 episomes and those from groups III,IV,V,VIII,IX and XI were missing both episomes. Retention of episomes upon reversion is characteristic of reversion under WF9 nuclear control. Loss of episomes upon reversion is characteristic of reversion under M825 nuclear control.
These results, although puzzling at first, reveal some aspects of the nuclear effect on plasmid retention. Although the nuclear dosage at the time of reversion in groups I, II, IV, VIII and X was about 50% WF9 and 50% M825, the genetic crosses performed to obtain the above mentioned revertants differed in each group. Revertants of group I and II arose from a cross of an RD-WF9 sterile plant with an M825 maintainer line. However, revertants of groups IV, VIII and X were obtained after crossing RD-WF9 sterile plants to the M825 maintainer line three (group IV), five (group VIII) and ten (group X) times, respectively, and then back to UF9 once.
Mitochondrial DNA of group I, II, III, IV, V, VI, VIII and X revertants has been subjected to Southern hybridization analyses using various mtDNA probes (Table 2). The patterns of mtDNA rearrangements in revertants of group I,II and X (50% WF9-50% M825) are equivalent to those observed by Escote et al. (Theor. Appl. Genet. 75:659-667, 1988) for cyto-revs in a 100% WF9 nuclear background or those we have observed in cyto-revs of group VI (89% WF9 nuclear background). The patterns of mtDNA rearrangements in revertants of group IV (56% WF9-44% M825), V (78% WF9-22% M825) and VIII (51.6% WF9-48% M825) are equivalent to those obtained by Escote et al. for cyto-revs in a 100% M825 nuclear background or those we have observed in cyto-revs of group III (87.5% M825 nuclear background).
Our results to date suggest that the nuclear effect on reversion is not a simple case of M825 dominance. At least one or possibly two backcrosses to M825 (see results of groups III and IV cyto-revs in Table 1) are required to change the WF9 reversion pattern to that of M825. Escote et al. (in preparation) have shown that, in the absence of reversion, mtDNA rearranges during the process of conversion of an RDWF9 sterile line to the M825 nuclear background. It is possible that a threshold level of these mtDNA rearrangements, mediated by exposure of the mitochondria to the M825 nucleus for two to three generations, needs to be attained before a reversion event can display the pattern of mtDNA reorganization characteristic of reversion in an M825 nuclear background. In other words, prior exposure of mitochondria to M825 nuclei alters their subsequent response to WF9 nuclei (see results of groups IV, IX and XI cyto-revs in Table 1). We have found an exception to this hypothesis in cyto-revs of group X. These revertants, obtained after crossing RD-WF9 sterile plants to the M825 maintainer line ten times and then back to WF9 once, retain the S1 and S2 episomes, a characteristic typical of reversion under WF9 nuclear control. Because of this nonconforming result obtained with group X cyto-revs, we have analyzed the hybridization patterns of several mtDNA probes to the sterile progenitor mtDNA from which revertants of group X were obtained (sterile RD-WF9/M82510) and an unrelated source of the same genotype. Interestingly enough, the hybridization pattern of the group X sterile source differed from the unrelated sterile source with pZmS4 ATPase 9 and IS1E5(R) probes. The observed mtDNA rearrangements in the sterile progenitor of group X cyto-revs could possibly be responsible for the aberrant mtDNA organization of group X revertants. We plan to examine additional cyto-revs derived from sources with the same genetic constitution as the one from which the group X revertants were obtained but unrelated to it. If the mtDNA reorganization of these revertants follows the pattern of reversion in an M825 nuclear background, we plan to investigate further the abnormalities of the sterile source of group X revertants which obviously may have something to do with the retention of S-episomes and the other mtDNA rearrangements characteristic of reversion under the influence of WF9 nuclear background.
Analysis of group VI revertants which arose after crossing an RDWF9
sterile plant to an M825 line three times and then back to a WF9 line three
times showed that these revertants retained the S episomes (characteristic
behavior of reversion in a WF9 nuclear background). This contrasts with
the results obtained with revertants of groups IV (RD-WF9/M8253xWF9) and
V (RD-WF9/M8253xWF92), which lose the episomes, suggesting that effects
such as the mtDNA rearrangements and possibly other changes imposed by
exposure to the M825 nucleus are reversible.
Return to the MNL 63 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}