--Susan L. Neuhausen
The Iowa Stiff Stalk Synthetic (BSSS) has been a source of successful inbreds and derivative lines. B14 and B37, from the initial cycle, and B73, from the fifth cycle, have proven useful as parents in commercial hybrids and as breeding germplasm in the development of recycled lines. Restriction fragment length polymorphisms (RFLPs) may be a useful tool to characterize this population at a genetic level. RFLP analysis was performed on the BSSS parents; the 4 inbreds comprising IA13, the tester for BSSS(HT); 12 inbreds released from BSSS cycles; 5 B14-derived inbreds; 2 B37-derived inbreds; 2 B73-derived inbreds; 2 testers for BSSS inbreds; 3 parents of some of the derived inbreds; and 124 individuals from the BSSS(HT)C5. The 170 lines were examined with 42 clones, and the 46 inbreds were examined with an additional 28 clones, spaced throughout the genome.
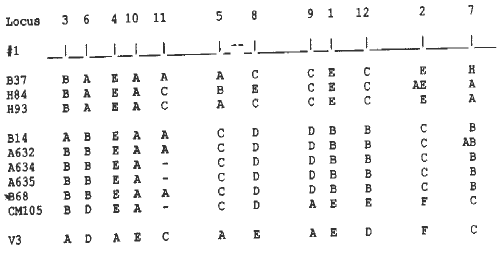
Conservation of chromosomal segments among derived inbreds and contributions from donor parents can be readily observed and followed using RFLPs. In Figure 1, alleles of inbreds are shown for loci on chromosome 1. H84 and H93 were highly conserved for B37 alleles except around the centromere and at the end of the short arm. There was strong conservation of B14 on chromosome 1 for all of the derived inbreds except for CM105, which had many alleles from V3, the donor parent, on the long arm.
Figure 1. Genotypes of inbreds for chromosome 1.
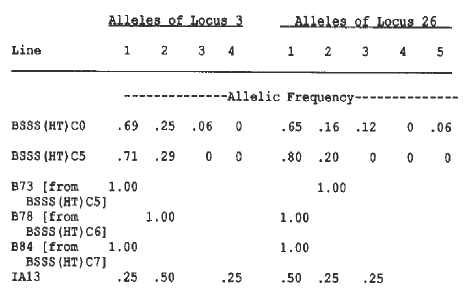
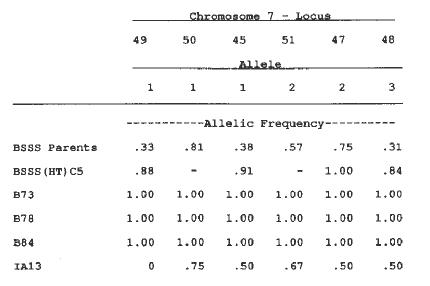
Changes in allelic frequencies from the original BSSS to the BSSS(HT)C5 may be due to selection for favorable alleles at those loci. In Table 1, two loci are shown as examples of little change in allelic frequencies from the BSSS(HT)C0 to the BSSS(HT)C5. This suggests that there was no selection pressure or genetic drift for these loci. In contrast, in Table 2, two loci are shown which exhibited large changes in allelic frequencies probably resulting from selection. Allele 3 for locus 4 increased from 0.19 to 0.83 in 5 cycles, such that B73 and B84 have that allele. Allele 4 for locus 4 was probably an important allele for that locus because its initial gene frequency was very low, yet it was maintained and was found in an inbred released from the sixth cycle. The allele was from CI.540. For locus 42, the allelic frequency of allele 1 increased and the three inbreds subsequently released contained that allele. Allele 2 for locus 42 decreased in the BSSS(HT) and was found in the tester. This is suggestive that there was a favorable heterotic response of allele 1 to the tester and no response for the additive effect of allele 2. On chromosome 7, gene frequencies changed from the BSSS(HT)C0 to the BSSS(HT)C5 such that all of the later cycle inbreds were of one allele type (Table 3), except for a locus at the end of the chromosome. This is a highly conserved chromosome for these inbreds due to gene frequencies approaching fixation. These results are suggestive that there is a selective advantage for these alleles at these loci and that selection pressure was on the entire chromosome. There were large shifts in gene frequencies from BSSS(HT)C0 to BSSS(HT)C5 for some of the loci and were likely due to effects of selection. A field evaluation of the BSSS(HT)C5 for yield in relation to IA13 and for stalk quality, with subsequent analysis for identification of QTLs, is necessary in order to confirm that changes were due to selection.
Table 1. Examples of little change in allelic frequencies from BSSS(HT)C0 to BSSS(HT)C5.
Table 2. Examples of allelic frequency changes from BSSS(HT)C0 to BSSS(HT)C5 likely due to selection.
Table 3. Loci on chromosome 7 at which one allele was conserved for inbreds released from later BSSS(HT) cycles.
It was difficult to assess whether individual BSSS parents were more
important than others in the success of this synthetic. Of the 402 alleles
examined for the 70 loci, only 91 alleles were represented by only one
inbred. Therefore, in most instances, a determination of which parent contributed
an allele was not possible. Only 28 of the 91 rare alleles were found in
the BSSS(HT)C1 inbreds, the BSSS(HT)C5, B73, B78, or B84. Many of the rare
alleles were probably lost due to drift. Of the 9 rare alleles which were
in at least one of the three inbreds released in the fifth (B73), sixth
(B78), or seventh (B84) cycles, two each were from CI.540, LE23, ILL.HY,
Ia.OS420, and one from Ind.B2. This is suggestive that these parents were
more important than others in the success of this synthetic.
Return to the MNL 63 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}