University of Minnesota
PHOENIX, ARIZONA
Desert Botanical Garden
--John Doebley and Gary Nabhan
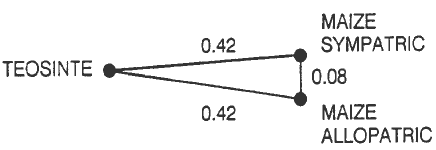
Hybrids between maize and its nearest wild relatives, the teosintes, are frequently observed where these plants occur sympatrically in Mexico and Central America. For this and other reasons, it has often been assumed that there is "constant gene flow" between maize and some types of teosinte. The alternative view that selective forces act to prevent substantial gene flow, especially from maize into teosinte, has been supported by one of us (J.D.) on the basis of field observations and evidence garnered from allozyme and chloroplast DNA studies. To further examine this question, one of us (G.N.) collected seed of (1) a teosinte (Z. mays subsp. mexicana) population from the Nobogame valley in Chihuahua, Mexico, (2) a maize landrace sympatric to this teosinte population (from the same field), and (3) a similar maize landrace allopatric to teosinte (from the nearby village, Baborigame) but in similar environmental setting. Thirteen to 16 individuals of each of these three collections were analyzed for their allozyme constitutions in accordance with procedures we have used previously; however, three new enzyme assays (adenylate kinase, hexose kinase and shikimic acid dehydrogenase) were also employed. Figure 1 shows the Rogers' distances between the collections. From this figure, it is clear that Nobogame teosinte is not genetically more similar to maize growing with it than it is to the same type of maize growing elsewhere, i.e., maize and teosinte growing in the same field and forming occasional hybrids do not become allozymically homogeneous. This conclusion is further supported by an examination of the actual allelic frequencies (Table 1), which show that teosinte maintains an allozymic constitution distinct from that of maize growing sympatrically with it. Of the 16 alleles that are found in only two of the three collections, 13 are shared by the two maize collections, two are shared by teosinte and the sympatric maize, and one is shared by teosinte and the allopatric maize.
Figure 1. Rogers' distances between a Nobogame teosinte population and maize growing sympatrically and allopatrically with it.
These results are in agreement with previous studies of the allozymic constitution of sympatric maize and teosinte populations. Taken together, these studies provide convincing evidence that, despite the formation of hybrids, maize and teosinte are under strong disruptive selection that (1) prevents the free flow of germplasm between them and (2) enables them to maintain distinct genetic constitutions for loci other than those controlling the differences in the morphology of their ears.
Table 1. Isozyme allele frequencies for a collection of Nobogame teosinte,
maize growing sympatrically with this teosinte and maize growing allopatrically
to this teosinte but from the same general region.
| Locus-allele1 | Maize alone | Teosinte | Maize with Teosinte | |
| Acp1-a | (2) | 0.731 | 1.000 | 0.737 |
| -b | (3 or 4) | 0.269 | 0.000 | 0.263 |
| Adh1-a | (4) | 1.000 | 1.000 | 1.000 |
| Adk1-a | 0.808 | 1.000 | 0.868 | |
| -b | 0.192 | 0.000 | 0.132 | |
| Enp1-a | (6 or 7) | 0.961 | 1.000 | 0.947 |
| -b | (8) | 0.039 | 0.000 | 0.053 |
| Glu1-a | (1) | 0.125 | 0.000 | 0.053 |
| -b | (?) | 0.042 | 0.000 | 0.053 |
| -c | (2) | 0.125 | 0.063 | 0.237 |
| -d | (3) | 0.042 | 0.000 | 0.000 |
| -e | (6) | 0.292 | 0.126 | 0.263 |
| -f | (7) | 0.208 | 0.000 | 0.211 |
| -g | (7.8) | 0.000 | 0.000 | 0.026 |
| -h | (8) | 0.000 | 0.813 | 0.053 |
| -i | (10) | 0.083 | 0.000 | 0.026 |
| -j | (12) | 0.000 | 0.000 | 0.079 |
| -k | (n) | 0.083 | 0.000 | 0.000 |
| Got1-a | (4) | 0.731 | 0.031 | 0.842 |
| -b | (5.8) | 0.000 | 0.969 | 0.000 |
| -c | (6) | 0.269 | 0.000 | 0.158 |
| Got2-a | (3) | 0.000 | 0.344 | 0.000 |
| -b | (4) | 0.961 | 0.656 | 1.000 |
| -c | (2) | 0.039 | 0.000 | 0.000 |
| Got3-a | (4) | 1.000 | 1.000 | 1.000 |
| Hex2-a | 0.115 | 0.094 | 0.079 | |
| -b | 0.885 | 0.906 | 0.921 | |
| Idh2-a | (4) | 0.500 | 0.438 | 0.500 |
| -b | (6) | 0.500 | 0.562 | 0.500 |
| Mdh1-a | (1) | 0.000 | 0.000 | 0.079 |
| -b | (6) | 1.000 | 0.000 | 0.920 |
| -c | (10.5) | 0.000 | 1.000 | 0.000 |
| Mdh2-a | (3) | 0.077 | 0.031 | 0.290 |
| -b | (3.5) | 0.115 | 0.000 | 0.000 |
| -c | (6) | 0.808 | 0.969 | 0.710 |
| Mdh3-a | (16) | 0.885 | 1.000 | 0.974 |
| -b | (18) | 0.115 | 0.000 | 0.026 |
| Mdh4-a | (12) | 1.000 | 1.000 | 1.000 |
| Mdh5-a | (12) | 1.000 | 0.219 | 0.921 |
| -b | (15) | 0.000 | 0.781 | 0.079 |
| Mmm1-M | (M) | 1.000 | 1.000 | 1.000 |
| Pgd1-a | (2) | 0.462 | 0.000 | 0.500 |
| -b | (3.8) | 0.538 | 1.000 | 0.500 |
| Pgd2-a | (5) | 1.000 | 1.000 | 1.000 |
| Pgm1-a | (9) | 0.807 | 0.208 | 0.900 |
| -b | (16) | 0.077 | 0.792 | 0.100 |
| -c | (?) | 0.115 | 0.000 | 0.000 |
| Pgm2-a | (3) | 0.077 | 0.042 | 0.079 |
| -b | (4) | 0.923 | 0.958 | 0.921 |
| Phi1-a | (2) | 0.115 | 0.000 | 0.079 |
| -b | (3) | 0.039 | 0.031 | 0.158 |
| -c | (4) | 0.846 | 0.969 | 0.763 |
| Sad1-a | 0.077 | 0.000 | 0.053 | |
| -b | 0.923 | 0.937 | 0.947 | |
| -c | 0.000 | 0.063 | 0.000 |
1The parenthetical allelic designations are those used by
C. Stuber and co-workers and are tentatively assigned.
Return to the MNL 63 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}