Punjab Agricultural University
--V. K. Chaudhary and J. L. Minocha
This investigation was designed to ascertain whether ethyl methane sulphonate (EMS), ethidium bromide (EB) and streptomycin (SM) can create cms mutants in maize. Seeds of inbred CM400 were soaked in a water solution of 0.1 and 0.2 percent EMS, EB and SM for 24, 48, and 72h at 5±1 C. Treatments of EMS (0.2%) and EB (0.3%) for 8h at 25±1 C were also used. Three male-sterile mutants were observed in M1 generation, i.e., one in the EB (0.2%) for 48h at 5 C and two in SM (0.1%) for 48h at 5 C treatments. These mutagens along with EMS (0.1% and 0.2%) also induced such male-sterile mutants in M2. Non-stainability of male-sterile pollen grains ranged from 85.7 to 99.2 percent as compared to the control (7.8%). It was difficult to distinguish visually between fertile (F) and sterile (S) plants of the same inbred line before flowering. The frequency of male-sterile mutants ranged from 7.5 to 26.6 (EMS), 1.2 to 23.2 (EB) and 2.8 to 44.5 (SM) on a 1000 plant basis for the M2 population of approximately 25,000 plants. To find the nature of the mutation induced, the male-sterile mutants were crossed with pollen from untreated inbred parent. The progenies of these mutants segregated in the subsequent generations up to M4 producing sterile as well as fertile plants. One of the progenies from EB treatment segregated to give male-sterile plants even in M5 generation. Occurrence of male-sterile mutants in some of the progenies right from M1/M2 up to M4/M5 generations clearly demonstrate that this trait is not controlled exclusively by nuclear genes (genetic). This conclusion further gets support from the study of segregation behaviour of (i) selfed male-fertile sibs of male-sterile progenies and (ii) male-fertile sibs in reciprocal crosses with the control. Some of these selfed and reciprocally crossed plants gave male-sterile mutants, giving an indication of the involvement of nuclear genes, and in some of the cases no male-sterile plants were observed, giving the indication that nuclear genes were not involved in the production of male-sterile mutants. There seems to be differential expression of the character under study.
In the past such attempts have also been made by Briggs (MNL 45:13-16, 47:35-37, 50:28-29, 51:5-6, 52:18-19) and Burnham (MNL 47:127-128), using different concentrations of EMS, EB and SM.
Male sterility induced in these progenies cannot be genetic because in such cases where the F1 of male-sterile x control (fertile) should possess plants with normal pollen fertility, several crosses segregating for male-sterility traits were found.
In this study it appears that the male-sterility induced could be attributed to the interaction of both cytoplasmic and nuclear genes. Since induced male-steriles and their fertile sibs segregated giving male-sterile as well as fertile individuals, it can be assumed that the genome carries some sort of partial restorer genes or modifiers. An attempt now may be made in the subsequent generations to try a number of inbreds for the purpose of maintaining this male-sterility. According to Singh and Laughnan (Genetics 71:607-620, 1972) it also appears at least equally plausible that the male-fertility encountered in the male-sterile progenies may be the result of an occasional contribution of normal cytoplasm by the male-fertile maintainer parents which are employed routinely as pollinator in crosses with male-sterile mutants. This needs to be meticulously investigated.
Morphological characterization. Mutants were characterized morphologically for spike characteristics, anther dehiscence and anther size.
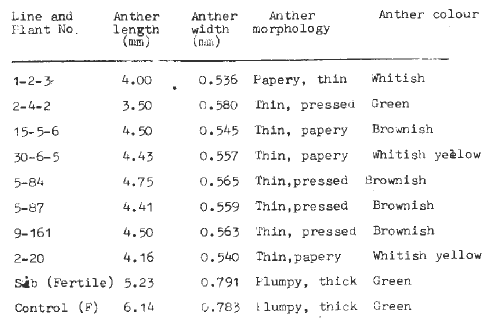
Spike characteristics - The inflorescence and the individual florets of male-sterile plants were rather smooth in touch and appearance. Maybe this is due to the non-plump nature of the anthers. The florets had unextruded anthers. Anthers when examined were observed to be thin, papery, small and pressed in shape and size and had different colours like green, whitish, and whitish yellow (Table 1). The matured florets of CM400 (control) on the other hand had extruded anthers and were thick, plump, green and bigger than those of the male-sterile mutant. For testing the spikelet fertility a few of the male-sterile mutants when selfed gave no seed set whereas seed set was normal when crossed to the control.
Anther dehiscence - There was no dehiscence of anthers in any of the male-sterile mutants as compared to the control, where there was complete dehiscence. This characteristic was used as one of the criteria to pick up male-sterile mutants in field conditions.
Table 1. Morphology, colour and size of anthers of male-sterile mutants and control.
Anther size - Anther length and width of male-sterile mutants were reduced as compared to the anthers of the control inbred parent (Table 1). The range of anther length and width was 3.50 to 4.75mm and 0.534 to 0.580mm respectively as compared to the control, where length ranged from 5.23 to 6.14mm and width from 0.783 to 0.791mm.
It was difficult to distinguish visually between fertile (F) and male-sterile (S) mutants of the same inbred line before flowering. However, at flowering tassels of male-sterile mutants and the control inbred differed in their degree of anther exsertion as discussed earlier.
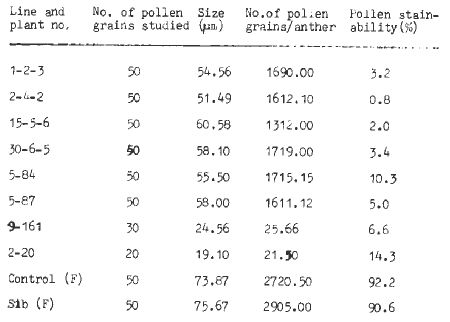
Cytological characterization. Pollen stainability (in 0.2% I-KI and 1% acetocarmine stain) was very low (below 15%) in the case of male-sterile mutants as compared to the control (Above 90%) inbreds (Table 2). Using ocular and stage micrometer, reduction in the pollen grain size (at the maximum dimension) was also observed in the case of male-sterile mutants when compared with the control. This size varied from 19.10 to 60.58µm in male-sterile mutants as compared to the control inbreds (73.87 to 75.67µm). On the basis of dehisced pollen grains per anther, two types of male-sterile mutants were observed. In one category (Type 2) there were abundant dehisced pollen grains per anther (1312 to 1719) while in the other (Type 1) there were few (11.00 to 25.66) pollen grains per anther as compared to the control inbred (2700-2905). This study was done by crushing the individual anther of male-sterile and control (fertile) plants on separate glass slides and mounting in 0.2 I-KI (stain) solution. A count of dehisced pollen grains was made and calculated per anther. It was ensured that no pollen grains went out of the field covered by the coverslip. This could be done by using a very small judicious amount of I-KI solution. Generally ten anthers from different spikelets were sampled from the male-sterile and fertile plants. The meiotic behaviour of male-sterile mutants (Type 2) was studied. The PMC's were examined at diakinesis, metaphase I, anaphase I and anaphase II. At least 100 cells from each of the mutants were studied. All mutants showed regular bivalent formation at diakinesis and metaphase I. There was normal separation of chromosomes at anaphase I and anaphase II. These male-sterile mutants were examined in M2 and later generations for meiotic chromosomal behaviour and no such abnormality has been observed so far.
Table 2. Pollen grain size and pollen grains/anther of male-sterile mutant and fertile plants.
Biochemical characterization. Quantitative as well as qualitative (isozyme) differences of some of the enzymes, viz. peroxidase, esterase, acid phosphatase and amylase (only isozyme) were studied. In the present study biochemical levels of soluble proteins, peroxidase, acid phosphatase and esterase were assayed in male-sterile mutants MS-1 and MS-3 (with few pollen grains in the anther), MS-2 (with many pollen grains), their fertile sibs (FS) and control (CF). All the assays were carried out from the mature anther extracts. In male-sterile mutants and their fertile sibs there was an observed decrease in the soluble protein level as compared to the control. The maximum reduction was in MS-1 followed by MS-3, MS-2 and fertile sibs. Specific activity of peroxidase was about nine and one half times more in MS-2 and fourteen times more in MS-1 and MS-3 as compared to the control. For acid phosphatase and esterase specific activities were less than the control in all the male-sterile mutants.
The isozyme patterns in mature anther extracts of male-sterile mutants obtained through starch gel electrophoretic studies were different from those of the control. In total, eleven isozymes of peroxidase (nine anodal and two cathodal), three of acid phosphatase (all anodal), six of esterase (all anodals) and five of amylase have been observed. The intensity and the number of bands of peroxidase isozyme were much higher in male-sterile mutant anther extracts than their fertile counterpart. However, isozymes of acid phosphatase and esterase showed higher intensity of bands in the control than in male-sterile mutant mature anther extracts. The results obtained from isozyme pattern clearly support the study of enzyme activities for peroxidase, esterase and acid phosphatase.
Isozyme pattern in the case of amylase showed more enhanced activity in extracts of mature anthers in male-sterile mutants than in the control.
The enzyme activities were calculated in terms of intensity scores from electrophoretic patterns of the enzymes. The results obtained were similar to the ones obtained by direct enzyme assay. This proved to be meaningful criteria for characterization of male-sterile mutants when comparing with fertile and normal plants.
These large numbers of isozymes for different enzyme systems indicate multiple gene control where there seems to be protein multiplicity in anthers of different male-sterile and fertile plants.
Histological characterization. Light microscopic comparisons of anther histology of fertile plants (control) and cms mutants at different stages of anther development revealed two distinct types of histological changes (Type 1 and Type 2) causing pollen abortion in cms mutants. The following disturbances were noted at different developmental stages in Type 1: (i) the development of vacuolate sporogenous cells was not normal as compared to the control; (ii) the tapetal layer was found to be separated from the sporogenous cells; (iii) due to non-secretion of callose, the sporogenous cells did not undergo meiosis to form tetrads and microspores, leading to the absence of pollen grain; and (iv) as a result, the entire locule bore an empty look with the remains of the sporogenous cells adhered to the internal wall of the locule in the streak form. Thus no pollen grains were formed. However, in Type 2, the development of the vacuolate sporogenous cells was normal at the callose stage but exhibited a gap between tapetum and endothecium contrary to the control. Splitting of sporogenous cells, formation of tetrads and microspores were normal. However, microspores released in the locule cavity were irregular in their outline due to the simultaneous breakdown of the tapetal layers, which have a nutritive role to play in the normal development of the pollen grains. The tapetal breakdown was further found to be accentuated in later stages, leading to the formation of disorganized, crumpled and irregular pollen grains thus causing pollen abortion. Thus, in Type 1 the disturbances in the pathway of anther development took place at callose stage due to non-splitting of sporogenous cells, whereas in Type 2 it started at the microspore stage leading to tapetal breakdown.
Effect of treatment temperature, duration and crop-season on the frequency of induced male-sterility. Eighteen treatments of EMS, EB and SM with 0.1 and 0.2 percent concentrations were used for 8, 24, 48 and 72 hrs. The subsequent generations were raised in summer (June-July) as well as winter (Oct.-Nov.) to advance the generations. Among these, only three (7th, 10th, and 13th) treatments were administered at 25±1 C and the rest at 5±1 C (Table 3).
Table 3. Mutation frequency in relation to treatment temperatures, duration and crop season. (Table not available - see hard copy.)
These three treatments produced male-sterile mutants in the frequency
of 26.6, 13.2, 4.4 respectively in winter (20±1 C) and 8.4, 18.7,
23.2 respectively in summer (30±1 C). The frequencies of 26.6 and
23.2 (Table 3) are apparently higher as compared to others in different
treatments carried out at 5 C except that of SM (44.5). In the case of
EMS (0.1%) there appears to be an increase in the frequency of male-sterile
mutants up to 48 hrs of treatment but not thereafter. After EB treatment
of 0.1% (4-6) there was no effect except the one given for 72 hrs. In EMS
(0.2%) treatment (7-9) there were no male-sterile mutants observed except
at 8 hrs in either season. An increase in mutation frequency was recorded
up to 48 hrs in the treatment of EB (0.2%) (10-12) and a decrease thereafter.
The frequency of male-sterile mutants in EMS treatment 7 (0.2%) was observed
as 26.6 and 8.4 and in EB treatment 10 (0.2%) it was 13.2 and 18.7 in winter
and summer respectively. There was no recovery of such mutants in the EMS
0.2% treatment (8-9) in either season. In the EB and SM treatment the frequency
of male-sterile mutants recorded as low in winter and high in summer, whereas
the trend was just the reverse in the case of EMS treatments. A similar
pattern of results was obtained in winter and summer of 1984, 1985, 1986
, and 1987.
Return to the MNL 63 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}