--Wayne Carlson
Studies by Lin (Genetics, 1979) and Carlson (Ann. Rev. Genetics, 1978) suggest that the B chromosome is not telocentric. They found that rearrangements at the B centromere which should delete an adjacent short arm have a genetic effect: they reduce the rate of nondisjunction at the second pollen mitosis. The findings provide some evidence for the existence of a B short arm. However, another explanation is possible. Perhaps part of the B centromere itself can be removed without disrupting the basic functioning of the centromere. This component of the centromere affects the rate of mitotic nondisjunction. To accommodate both ideas, the B centromere plus the hypothetical short arm are referred to as region 4. Within this region, there is a site(s) which affects the rate of nondisjunction.
One explanation for the effect of region 4 is to assume that the active site is heterochromatin in the B short arm. The heterochromatin is similar to that in region 3, adjacent to the centromere in the long arm. Region 3 heterochromatin controls nondisjunction through non-division at the second pollen mitosis (Rhoades and Dempsey, Genetics, 1972; Carlson and Chou, Genetics, 1981). Perhaps maximal nondisjunctional rates require that the B centromere be flanked on both sides by non-dividing heterochromatin.
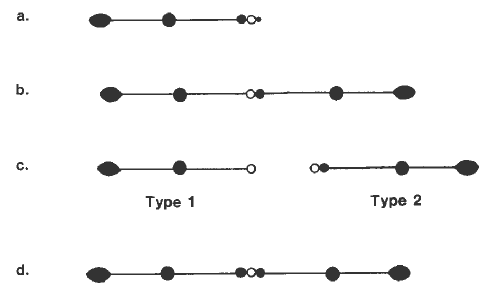
A test can be made for equivalency of region 3 and region 4. In the accompanying diagram, misdivision products of the B9 chromosome from TB-9Sb are shown. The following are depicted: a) standard B9, b) pseudoisochromosome derivative of B9, c) type 1 and type 2 derivatives of the pseudoisochromosome, d) type 2 isochromosome derivative of the type 2 telocentric.
The type 2 isochromosome contains two doses of region 3, flanking the centromere. It should give nondisjunctional rates similar to the standard B9, if region 3 is equivalent to region 4. An ear was constructed which segregated two types of progeny: 9B (Wx) 9B (Wx) type 2-iso B9 (Yg2 Bz) and 9 (wx) 9B (Wx) standard B9 (Yg2 Bz). Seven plants of each type were crossed as male to a tester of B9 nondisjunction: yg2/+ bz bz wx wx (only Wx kernels were used in the analysis, because they contain TB-9Sb).
Nondisjunctional rates were calculated as bz Yg plus 2 X Bz
yg2 phenotypes (the yg2 totals must be doubled due to heterozygosity
of the tester). Rates of nondisjunction for two ears per cross are given
below:
| 9B 9B type 2-iso B | |
| Plant No. | Rate Nondisjunction |
| 6072-1 | 46% (292/638) |
| -10 | 74% (532/718) |
| -28 | 43% (280/652) |
| -45 | 54% (302/559) |
| -50 | 49% (285/587) |
| -56 | ss% (399/723) |
| -70 | 57% (330/582) |
| 9 9B std B9 | |
| 6073-3 | 92% (338/369) |
| -6 | 92% (416/453) |
| -17 | 94% (361/385) |
| -21 | 90% (316/353) |
| -22 | 91% (364/399) |
| -23 | 99% (463/466) |
| -24 | 95% (362/382) |
There is a large, nonoverlapping difference in rates of nondisjunction
between the two groups. It is concluded that region 4 contains a unique
site of nondisjunctional control which cannot be replaced by nondividing
heterochromatin.
Return to the MNL 63 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}