University of Oregon
--Vicki L. Chandler, Devon Turks, and Pablo Radicella
The synthesis of anthocyanin pigments in maize plant tissues or seeds requires the products of numerous genes, both structural and regulatory (Coe and Neuffer, Corn and Corn Improvement, 1977). The R locus (on chromosome 10) regulates the formation and distribution of anthocyanin pigments in certain tissues of the plant and seed. B (on chromosome 2) controls anthocyanin formation in plant tissues independent of R, and several B alleles can substitute for R function in the seed. For example, B-Peru can substitute for R-g function in the seed, and only one functional allele at either locus is required for pigment synthesis. Experiments have demonstrated that both genes act on the same biosynthetic pathway in that a functional R or B allele is required for the presence of the structural enzyme encoded by the Bz1 gene (Dooner, Mol. Gen. Genet. 189:136, 1983; Gerats et al., Biochem. Genet. 22:1161, 1984).
We reasoned that the fact that certain B and R alleles are functionally duplicate may reflect some DNA sequence conservation between the two genes that might be detectable using Southern blot hybridization. To investigate this we have used the cloned R gene as a hybridization probe to screen Southern blots containing DNA samples prepared from various B alleles for restriction fragment length polymorphisms.
The 5.0kb R clone corresponding to the seed component (S) of R was obtained from S. Dellaporta. This is an R allele that is functionally duplicate to B-Peru. The clone obtained from Dellaporta contained a sequence highly repeated in the maize genome, so subcloning was required to obtain a relatively unique copy region to use on Southern blots. A variety of restriction enzymes and hybridization conditions were then utilized to detect a restriction fragment polymorphism among our various B alleles.
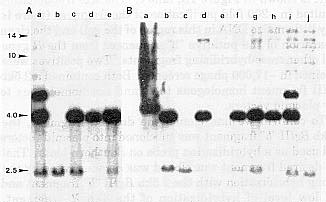
An example of one Southern blot showing a BglII restriction fragment polymorphism between several B alleles is shown in Figure 1A. The R allele (r-g) is constant in all of our B stocks and generates a 4.0kb BglII fragment. Our standard b tester stock contains a weakly hybridizing 2.5kb BglII fragment (lane c) which is not in the progenitor B-Peru stock (lane d). Instead we see a large ~14kb BglII fragment in the B-Peru stock. This ~14kb BglII fragment is missing in one of our B-Peru mutants (b-Perumu5) that contains a transposable element insertion into the B-Peru gene, and a ~10kb BglII restriction fragment is observed (lane e). (The b-Perumu5 individual with the insertion mutation is heterozygous for the b tester allele and contains the 2.5kb BglII fragment.) This is the pattern that would be expected if the fragments hybridizing to the R probe were linked to the various B alleles.
Figure 1. Southern blots on DNA from B and R stocks hybridized with the R probe. DNA samples were digested with BglII, electrophoresed on 0.5% agarose, transferred to nylon membrane (MSI), and hybridized with a 1.9kb R clone labelled by random hexamer priming. A: lanes a-e, DNA samples from maize stocks with the designated B and R alleles: a) b (null) R-g; b) b R deletion (DNA sample courtesy of Mary Alleman); c) b r-g; d) B-Peru, r-g; e) b-Perumu5/b, r-g. B: lane a) reconstruction containing ~1 copy per haploid genome of the ~5.0kb R clone (courtesy of S. Dellaporta); b) b r-g stock; c) size- fractionated DNA from b r-g stock; d) B-Peru r-g; e) size-fractionated DNA from B-Peru r-g stock; f) b-Perumu5/b r-g plant, g) b-Perumu5/b r-g plant; h) b-Perumu5/b-Perumu5 r-g plant; i) b R-g plant; j) b R deletion.
To further test this hypothesis additional Southern blots were performed on DNA samples from plants segregating the various B alleles. The larger BglII fragments observed in the B-Peru stock and the insertion mutant were difficult to reproducibly detect on the Southern blots, presumably due to the weak signal and poor transfer of large fragments on blots. We also frequently observed additional restriction fragments hybridizing to the R probe in this region of the gel, masking the fragments we were trying to follow (Figure 1B). Therefore, we decided to address whether the 2.5kb BglII fragment segregated with the b allele. We prepared DNA from plants that had been previously shown to be either b/b, b-Perumu5/b, or b-Perumu5/b-Perumu5. The ten individuals that were b/b or b-Perumu5/b all contained the 2.5kb BglII fragment, and none of the twenty-seven individuals homozygous for b-Perumu5 contained the 2.5kb BglII fragment. This result demonstrated that the 2.5kb BglII fragment is linked to the b allele. An example of several individuals is shown in Figure 1A, lanes c and e, and Figure 1B, lanes f-h.
The 2.5kb BglII fragment was cloned from the standard b tester stock, by size fractionating the DNA, cloning in a lambda vector that will carry 0-12kb BglII fragments (Bv2, obtained from Noreen Murray), and screening the resulting phage with the R probe using the conditions established with the Southern blots. An example of the size fractionated DNA is shown in Figure 1B, lane c. The size fractionation resulted in ~200 fold purification of the fragment (there is very little maize DNA in this region of the gel) and the clean separation of the putative "b" sequences from the R gene and other cross-hybridizing fragments. Two positives were obtained in ~17,000 phage screened. Both contained a 2.5kb BglII fragment homologous to R and not homologous to any plasmid vectors.
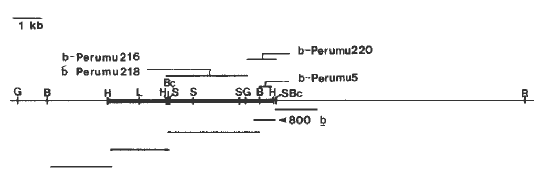
To confirm we had cloned the desired fragment, the 2.5kb BglII "b" fragment was subcloned into plasmid vectors and used as a hybridization probe on Southern blots. That the desired fragment was cloned was demonstrated by the strong hybridization with the 2.5kb BglII "b" fragment and the low level of hybridization of the 4kb R fragment. However, the 2.5kb BglII "b" fragment contained some repeated sequences, and further subcloning was required to obtain a relatively unique sequence. Using an ~800 bp BglI/HindIII subclone as a hybridization probe (see Figure 3), we have tested whether it recognizes the functional B gene. As described last year, we have isolated four independent transposable element insertion mutations into the B-Peru allele and revertant derivatives of each. These independent insertion mutants have been compared to the progenitor B-Peru allele utilizing Southern blots hybridized with the putative "b" sequences. An example of one Southern blot is shown in Figure 2. Each DNA sample was digested with BclI, which produces a 4kb fragment in the b tester and the progenitor B-Peru allele. Each of the mutants (b-Perumu5, b-Perumu216, b-Perumu218, and b-Perumu220) contain restriction fragment differences relative to the wild type B-Peru allele, suggesting that each mutant contains an alteration within this 4kb fragment. The "new" restriction fragments are indicated by arrows on the autoradiograph shown in Figure 2. Most importantly, in the b-Perumu216- revertant (lane g), the restriction fragment characteristic of b-Perumu216 is missing and only the progenitor 4kb fragment is observed. These results definitively demonstrate that the sequences we have cloned recognize the functional B-Peru gene. Further restriction mapping indicates that the restriction fragment variation between the different mutant alleles reflects a combination of where the element is inserted within the gene and different BclI sites within the various elements. Each insertion contains at least one BclI site. The 4kb BclI fragment is from the b allele, heterozygous in each mutant plant. The b-Perumu5 allele contains an insertion within the region spanned by the 800 bp probe, so that both halves of the element are observed. The other alleles contain insertions to the left of the 800 bp probe (as drawn on the map in Figure 3), so only one-half of each element is observed in this hybridization.
Figure 2. Southern blot on DNA samples from B and R stocks hybridized with the 800bp b sequences. DNA samples (~4ug) were digested with BclI, electrophoresed in 0.5% agarose, transferred to nylon membrane (MSI), and hybridized with the 800bp b fragment labeled by random hexamer priming. Lanes a-k, DNA from maize plants containing the designated B and R alleles: a) b r-g; b) B-Peru r-g; c) b-Perumu5/b r-g; d) b-Perumu5/b r-g; e) b-Perumu216/b r-g; f) b-Perumu216/b r-g; g) b-Perumu216-revertant/b r-g; h) b-Perumu218/b r-g; i) b-Perumu218/b r-g, the b allele in this stock is polymorphic relative to the other b allele shown; j) b-Perumu220/b r-g; k) b-Perumu220/b r-g. The b-Perumu5, b-Perumu216, b-Perumu218 and b-Perumu220 alleles are independent Mu-induced insertions into the B-Peru gene.
Currently, the b sequences are being used to clone the progenitor B-Peru allele and each insertion mutant allele, and to generate more extensive restriction maps of the various alleles. A restriction map summarizing our current knowledge of the B-Peru allele and where the insertions have occurred is shown in Figure 3. Our best estimate for insertion size within the various alleles is: b-Perumu5, 6.5kb; b-Perumu216, 3.0kb; b-Perumu218, 7.5kb; and b-Perumu220, undetermined. Various restriction fragments of the b and B-Peru alleles have been used on Northern blots to identify an ~2.2kb mRNA. The restriction fragments hybridizing to this transcript are indicated in Figure 3.
Figure 3. Restriction map of B-Peru allele, showing regions transcribed and sites of insertion. The restriction map was generated from a combination of genomic Southern blots, and digestions with cloned DNA sequences. The fragments indicated below the map are subclones used as hybridization probes to generate the restriction map and analyze transcripts on Northern blots. Abbreviations: G, BglII; B, BamHI; H, HindIII; Bc, BclI; L, SalI; S, SacI. The thickened restriction fragments hybridize to an ~2.2kb mRNA in B-Peru and B containing stocks. The various Mu-induced alleles contain insertions in the fragments, as indicated above the map.
The laboratories of Ben Burr and Steve Dellaporta have independently
cloned the B genomic sequences using a similar strategy, and our
results are consistent with the results of those laboratories.
Return to the MNL 63 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}