USDA-ARS and University of Missouri
BOMBAY, INDIA
Tata Institute of Fundamental Research
--M. M. Johri and E. H. Coe
We are examining the pattern of sectors in plants with the following factors affecting the chloroplasts: f (fine stripe); j1 (japonica-1); sr1, sr2, sr3 (striate-1, 2 and 3 respectively) and v5 (virescent-5). At the time of examination in August 1988, the pedigrees of the test material varied considerably (Table 1) and these factors are being crossed to ensure uniform background. Though the top 12 or 13 leaves have been examined, the plants exhibiting the sectors were identifiable early at the seedling stage.
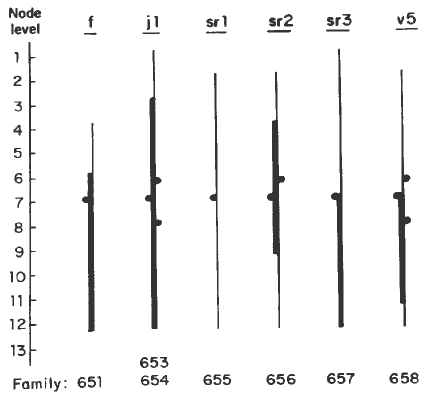
The expression of sectors at different nodes for various factors is shown in the Figure. Whereas fine stripe sectors were absent from the top 2 or 3 leaves, sectors for other factors were usually present in the top 12 or 13 leaves. In most cases, characteristically, the widest sectors were located along the edges of the blade, especially in the upper leaves. In the case of f, sr3 and v5, sectors were also present in the lamina. In sr1 plants, most of the sectors, including those along the edges of the blade, were narrow (1/16-1/32). A similar pattern is present in sr2 plants also, but in all cases the sectors along the edges in the leaf sheath were wider than those in the leaf blade. The extents of lateral proliferation of a lineage along the edges of sheath and blade show no relationship and these must be regulated by independent factors or genes. Since at least one cell lineage from sheath (along the margin) is known not to be represented in the leaf blade, the wide sectors at the margin of the sheath in sr2 could be derived from more than one clone.
Figure. Extent of striping at different node levels, counting from the top. Thick line represents maximum expression for that factor.
Table 1. Pattern of striping in several mutant types. [Table not available--see hard copy]
The abaxial surface of leaves in j1, sr3, and v5
plants was pale green indicating a displacement of LII-derived layers by
LI. The overall pattern of sectors suggests that the shoot apex of plants
(at least in j1, sr3 and v5) is chimeric for LI. Whether
the exclusive expression of these factors in LI depends on the specific
genes or on the background of the material is yet to be established. During
embryogenesis in corn, the shoot meristem is derived from the derivatives
of at least two cell layers (Poethig et al., Dev. Biol. 117:392, 1986).
The two cell layers of the shoot apex maintain more or less their distinctiveness
as most of the cell divisions are anticlinal. The periclinal divisions
occur during organogenesis and since such divisions occur randomly in space
and time, different types of variegation patterns are formed in plants
with a chimeric LI. It is of considerable interest to determine if genetic
factors regulate the time and location of periclinal cell divisions. The
presence of wide or small sectors could be genetically regulated.
Return to the MNL 63 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}