--J.D. Smith and B.G. Cobb
The inhibition of anthocyanin synthesis by vp/vp in A C R aleurone tissue is well known. Other known functions of vp/vp, e.g., failure to induce embryo dormancy, suppress lipase, suppress peroxidases, etc., are mimicked by the abscisic acid (ABA)-deficient mutants, e.g., vp2, vp5, etc., and both ABA and the Vp gene product are required to regulate these events during normal kernel development. However, all homozygous ABA-deficient mutants accumulate anthocyanins in A C R aleurone tissue. Thus, Vp regulation of anthocyanin synthesis has been presumed to be independent of ABA.
ABA-deficient mutants, except vp8, block carotenoid synthesis which facilitates early identification of homozygous kernels on segregating ears. Since ABA is derived from an unidentified xanthophyll, these kernels are not able to synthesize ABA, but they do contain ABA translocated from the heterozygous maternal plant. At 15 days after pollination (DAP), ABA-deficient embryos contain about 50% as much ABA as normal ones (MNL 61:39).
Since the regulation of lipase activity by ABA is concentration dependent (MNL 62:43), it seemed possible that induction of anthocyanin synthesis might be similarly regulated. If so, the ABA threshold level must be lower than the maternal ABA contribution. To test this, we harvested homozygous A C R ears at 5 DAP and cultured kernel blocks on media ± ABA ± fluridone (a chemical inhibitor of carotenoid and ABA synthesis).
Figure 1 shows kernel blocks at different ages cultured on the 4 media. Rows 1 to 4 were cultured on normal medium, + fluridone, +ABA and +fluridone +ABA, respectively. The columns show blocks at different ages, starting at 15 DAP on the left and increasing at 2 day intervals to 23 DAP on the right.
Although the differences in anthocyanin pigmentation are more dramatic in color, Fig. 1 (row 1) shows that A C R kernels on normal medium are colorless at 15 DAP, but color development increases with age. If ABA is added to the medium (row 3), anthocyanin accumulation is enhanced at all ages.
ABA levels in embryos cultured on media containing fluridone (100mg/l) are negligible (ca. 6 fmol/embryo at 15 DAP), and these levels decrease with age as the source of this ABA appears to be preformed carotenoids in the cob tissue of the kernel block at the time the ear was harvested (MNL 61:39). Fig. 1 (row 2) shows that kernels are white for the entire age sequence, indicating that both xanthophyll and anthocyanin synthesis are inhibited in kernels cultured with fluridone.
If both fluridone and ABA (10-4) mol) are added to the medium (Fig. 1 row 4), the kernels are white due to inhibition of carotenogenesis, but anthocyanins accumulate at essentially the same rate as on medium +ABA (row 2) at all ages. This indicates that ABA is essential for the induction of anthocyanin synthesis.
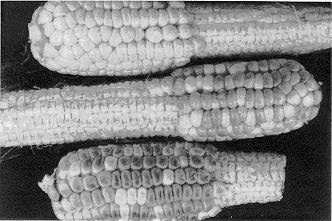
Figure 2 illustrates the importance of serendipity to science. These vp/+ ears were self-pollinated at Homestead, FL, on Nov. 19, harvested without husking on Dec. 14 and received in College Station, TX on Dec. 16 at 27 DAP. Surprisingly, these ears were colorless when husked, since vp/vp colorless kernels can be identified at 13 to 15 DAP in segregating ears developing at College Station in June. In Fig. 2, the dark colored ear was husked at 11:30 am, Dec. 16, the middle ear was husked at 11:30 pm, Dec. 16, and the colorless ear was husked at 11:30 am, Dec. 17. The ears were left on a lab bench after husking, and this photograph was taken shortly after the colorless ear was husked. Thus, these ears were allowed to desiccate at room temperature (72 F) for 0, 12 and 24 hours after husking. That desiccation occurred is illustrated by the noticeable denting of kernels 24 hours after husking.
Our interpretation of this is questionable, since we have not yet assayed these ears for ABA. However, we do know that ABA is stress induced and that ABA levels increase about 10-fold in maize seedling leaf tissue treated in the same way for one hour. Thus, we presume that mild temperatures in December and irrigation every third day created a non-stressful environment in the Florida nursery which resulted in abnormally low ABA levels in the plants and developing kernels, and these ABA levels were too low to induce anthocyanin synthesis. We further presume that ABA levels in aleurone tissue increased at a relatively rapid, though as yet undetermined, rate, since faint anthocyanin pigmentation was observed 5 hours after ears were husked.
Based upon these data, and similar data related to lipase, peroxidases and other enzymes, we have developed a working model which depicts how Vp and ABA may regulate events that occur during maize kernel development. This assumes that Vp codes for a soluble protein (VP) which may bind to upstream regions of specific genes. VP acts like a digital-type (on/off) transcriptional regulator. However, VP fails to function in the absence of ABA. Thus, ABA appears to function as an analog-type (concentration dependent) regulator of VP functions. ABA levels, in turn, appear to be regulated by external environmental conditions.
This may represent an example of environmental regulation of phytohormone
levels (ABA) which regulate the functions of a regulatory protein (VP)
which regulates transcription of specific genes during maize kernel development.
Return to the MNL 63 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}