Iowa State University
--James H. Zhou and Alan G. Atherly
The termini of Ac have prokaryotic promoters. The mechanism of transposition of plant transposable elements is not known, but the termini of the elements likely play an important role. We have found that an approximately 480bp DNA fragment of the 3' terminus of the maize transposable element Activator (Ac), when fused to the promoter-less gene encoding chloramphenicol acetyltransferase (CAT), turns on the CAT gene in E. coli strain DH5. Similarly, an approximately 178bp DNA fragment of the 5' end of Ac shows identical polarity and promoter activity as the 3' end. Both fragments are orientation-dependent in their promoter activity.
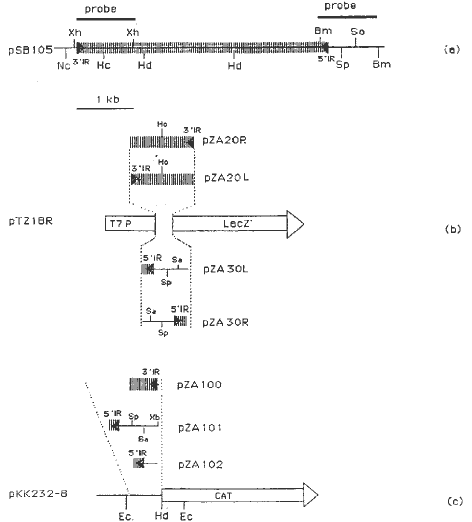
We inserted an approximately 1kb XhoI 3' end fragment of Ac element from pSB105 (Fig. 1) into vector pTZ18R at a SalI site within the multiple cloning sequence between the T7 promoter and the alpha-complementation fragment of the ß-galactosidase gene (LacZ') in two different orientations, giving rise to pZA20R and pZA20L. Similarly, the approximately 850bp 3' BamHI fragment of pSB105, containing the 178bp 5' end of Ac element was inserted into the modified pTZ18R (pTZ18R') BamHI site in two orientations, giving rise to pZA30R and pZA30L (Figure 1b).
The approximately 500bp HincII\ HindIII fragment of pZA20R, containing the 3' end of Ac, and the 850bp SmaI\ HindIII fragment of pZA30L, containing the 5' end of the Ac element, were each inserted into pKK232-8 (at the SmaI and HindIII sites, respectively) giving rise to pZA100 and pZA101, respectively. An approximately 650bp deletion was made in pZA101 (between the SphI site and XbaI site) to delete a portion of the flanking P-vv sequence from the 5' Ac end. This gave rise to pZA102 (Fig. 1c).
Fig.
1. Plasmid construction. a) The Ac fragment of P-vv present
in pSB105. The position of Ac is shown as the vertical-hatched bar
(same in b and c). The solid triangles indicate the direction of the inverted
repeat sequences (IR) of Ac (same in b and c). The thin horizontal
line extending out of Ac borders represents P-vv flanking
sequences (same in b and c). The solid bars above the Ac element
are the sequences used as probes of Ac termini in Figure 2. b) Schematic
representation of pZA20R, pZA20L, pZA30L and pZA30R plasmids. Insertion
of approximate 1kb XhoI fragment contained 3' end of Ac element
with only 22bp P-vv flanking sequence in either orientation and
insertion of approximate 0.85kb BamHI fragment contained 5' end
of Ac element in either orientation into the LacZ' gene in
plasmid pTZ18R are shown. The white triangle represents the direction of
LacZ'
gene expression. c) Schematic representation of pZA100, pZA101 and pZA102
plasmids. Insertion of approximate 480bp fragment of PZA20R (see b) containing
the 3' end of the Ac element with the 22bp P-vv flanking
sequence, and insertion of approximate 850bp fragment of pZA30L (see b.)
containing the 178bp 5' end of Ac element with the 662bp
P-vv
flanking sequence into the front of CAT gene. The pZA102 plasmid is derived
from pZA101 in which the approximately 650bp fragment of P-vv flanking
sequence is deleted at the SphI site and XbaI site between
the 5' end of Ac element and the 5' end of the CAT gene in pZA101.
Restriction endonuclease
abbreviations: Nc, NcoI; Xh, XhoI; Hc, HincII; Hd,
HindIII;
Bm, BamHI; Sa, SalI; Sp, SphI; Xb,
XbhI; Ec,
EcoRI.
Expression of the CAT gene was determined by growing transformed E. coli DH5 cells on LB plates containing 30mg\l chloramphenicol.
Expression of LacZ' gene is dependent upon the orientation of the inserted Ac termini. In pZA20R the direction of 3' inverted repeat sequence (3'IR) of Ac is opposite to that of the lacZ' gene; and in pZA20L the direction of 3'IR of Ac is the same as that of lacZ' gene. pZA20R shows lacZ' gene activity (Fig. 1). In contrast, pZA20L does not show any lacZ' activity. In addition, when the BamHI fragment of pSB105, containing the 5' end of Ac, was inserted into the BamHI site of the multiple cloning sequence of the vector (pTZ18R), in the direction of IR opposite to lacZ' gene (see Figure 1b: pZA30L) gene expression was observed. In the opposite orientation, no gene expression was observed. These experiments suggest that the termini of the Ac element have polar effects on expression of lacZ' gene that is analogous to that of prokaryotic transposons.
CAT gene expression driven by the termini of Ac. The Ac termini-CAT gene fusion is expressed when present in E. coli strain DH5. These strains can grow on agar media containing chloramphenicol at a concentration of 30mg/l. The negative control (pKK232-8 in E. coli DH5) cannot grow, even at 10mg\l chloramphenicol. These experiments indicate that the termini of the Ac transposable element have strong orientation-dependent prokaryotic promoter activities.
At least four putative prokaryotic promoter consensus sequences appear in a 500bp sequence within the 3' terminus (see Figure 2), and one in the 5' end of the Ac element, both within the "extended inverted repeat" region and close to the 11bp inverted repeat (IR) (the 5' sequence is not shown). We cannot be sure which putative promoter sequence in the 3' end turns on the CAT gene activity. However, since the putative promoter sequence in the 5' end is close to the perfect inverted repeat sequence (IR), we can reasonably speculate that the putative promoter sequence in the 3' end closest to the IR may turn on the CAT gene activity in pZA100. Thus, the two putative promoters in each end are likely located within the first mismatched "loop" of the secondary structure at the termini of the Ac element.
Fig. 2. The sequence of 500bp of the 3' end of Ac is presented. The numbers correspond to the position on the Ac sequence; the 3' correlates to the 3' of the 3.5kb transcript of Ac. The putative -10 and -35 consensus sequences are boxed. The ARS sequences are under-dotted. The possible start points (ATG) for transcription are underlined. The over-dotted sequences are homologous sequences with dnaA protein binding sites.
The probability by chance alone of a putative promoter consensus sequence residing in the 3' end of Ac element (pZA100) is approximately 1x10-4. Similarly, the probability of a promoter sequence in the 5' end is approximately 4x10-5. However this calculation assumes an equal distribution of the four bases in the sequences. These calculations suggest that it is unlikely for the promoter consensus sequences to appear by chance alone.
Sequence similarities between the 3' end region of Ac and oriC. A remarkable feature of the nucleotide sequence of the 3' end of the Ac transposable element is the abundance of TTATACACA sequences (four within a 250bp stretch, Fig. 2). This feature is reminiscent of the oriC region in which four consensus TTAT[C\A]CA[C\A]A sequences are present within a 245bp sequence and constitute the minimal origin of replication. These four inverted consensus repeats have been found to be DnaA protein binding sites. The DnaA protein is essential for DNA replication. Experiments with the 3' end of Ac as an origin of replication (XhoI fragment of pSB105 linked to a Kanamycin resistance gene, Fig. 1a) demonstrated that it was unable to function as an origin of replication when present in E. coli DH5 (data not shown). This could be due to several reasons: orientation of the binding sequences (direct repeats in Ac end and inverted repeats in oriC), the match with the DnaA protein complex (#2 is matched, but the remainder are not perfectly matched with the consensus sequence in oriC), lack of enough GATC methylation sites, or lack of "13 mer" sequences which are necessary for oriC function.
The 3' terminus of Ac possesses an ARS sequence. Examination of the sequence of the 3' terminus of the Ac element revealed the presence of two 11-mers consisting of the consensus sequence of the yeast ARS sequence (Figure 2). The 460bp 3' terminus of Ac was placed in a pUC18 vector lacking a yeast origin of replication (pUC-ura3,) creating pZA400. This vector possessed a ura3 gene for selection and was transformed into a ura3- yeast strain, selecting for growth in the absence of uracil. Controls included a similar vector with an ARS and one lacking ARS sequences. Transformants possessing pZA400 grew normally on uracil minus media, and segregated out when placed in media lacking uracil. These data strongly suggest that the 3' terminus of Ac possesses a functional ARS sequence.
An important question that arises from this study is how were the prokaryotic promoter-like sequences, DnaA protein binding site-like sequences, and ARS sequences in the termini of Ac element generated, and what functions do they exert? As suggested from the data of others and our own data, Ac termini when placed adjacent to promoter-less genes do not express activity in transformed plant cells. Our observations suggest that plant transposable element systems and prokaryotic transposons may share some similar "DNA signals" relating to the origin of DNA replication. This implies that the origin of the Ac transposable element family may have an evolutionary relationship with prokaryotic DNA replication origins and/or transposable elements. An attractive explanation for the presence of prokaryotic promoter sequences, ARS sequences, and DnaA protein binding site-like repeat sequences within the termini of Ac is that they are conserved as secondary sequences required for transposition during evolution. Evidence to support this hypothesis is that TTATACACA sequence is found in TRL-Tn5 and dnaA mutations affect the transposition efficiency of Tn5. Alternatively, additional factors may be required for transposition of DNA elements in plants. This could be a protein which recognizes the homologous sites of the prokaryotic DnaA binding sequence. It is possible that DnaA protein-DnaA-box interaction plays an important role in transposition, or regulation of transposition, as suggested from findings with bacteria.
We are presently conducting deletion analysis of the 3' terminus to
ascertain exactly which of these sequences are required for transposition.
Return to the MNL 63 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}